II. 39. Intragenic recombination at the waxy locus in Hordeum vulgare.
J. Rosichan, R. A. Nilan, P. Arenaz and A. Kleinhofs, Department of Agronomy and Soils and Program in Genetics, Washington State University, Pullman, Washington 99164, U.S.A.
The kinds of mutations, e.g., deletions, base substitutions, additions, induced by chemical or physical mutagens are not usually determined in eukaryotes. Systems for high resolution genetic analyses of the mutants, and biochemical analyses of the mutant gene product, are generally not available. This is a major gap in assessing the hazards of environmental mutagens.
The waxy locus of barley which controls starch composition is ideally suited for the development of a highly sensitive test system for monitoring and screening mutagens in man's environment. Forward and reverse mutations and mutant recombination frequencies within the waxy gene can be measured on a per million pollen grain basis--thus providing extremely high genetic resolution, a detailed map of the gene, and knowledge of the nature of the mutation events.
A procedure was developed to screen genetic changes at the waxy locus in Hordeum vulgare using analysis of male gametophytes. This method was adopted from procedures for investigating intragenic recombination in maize at the waxy (1) and alcohol dehydrogenase (2) loci. Pollen grain starch was stained with a solution of KI-I2. The stained pollen were vacuum deposited onto gridded Millipore membrane filters, counted, and screened for starchy pollen. The size of a pollen population was estimated by counting the number of pollen grains in a 0.1 ml aliquot from a pollen suspension. The number of pollen grains counted was multiplied by the volume and dilution to obtain the total number of pollen grains in the suspension.
It is possible to routinely and accurately screen large numbers of pollen grains to monitor low frequency genetic events at this locus. Forward mutations (blue staining -> red staining pollen), reverse mutations and allelic recombinants (red staining -> blue staining pollen) can be distinguished by this procedure.
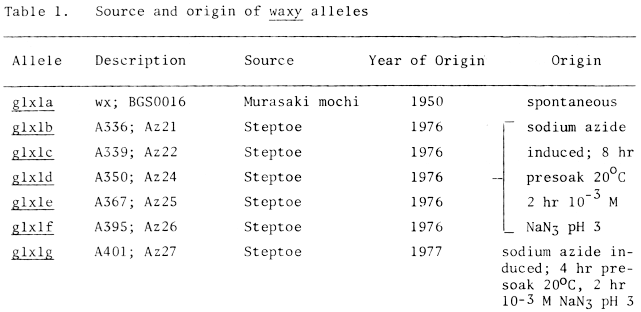
The barley (Hordeum vulgare) cultivar 'Murasaki mochi' was the source of the standard waxy allele. Six other waxy alleles analyzed in this study were induced in the cultivar 'Steptoe' by sodium azide and were selected by a procedure reported previously (3). Their origins are outlined in Table 1. Previous results (unpublished data) indicated that all six azide-induced waxy alleles and the standard waxy allele are allelic.
Table 1. Source and origin of waxy alleles
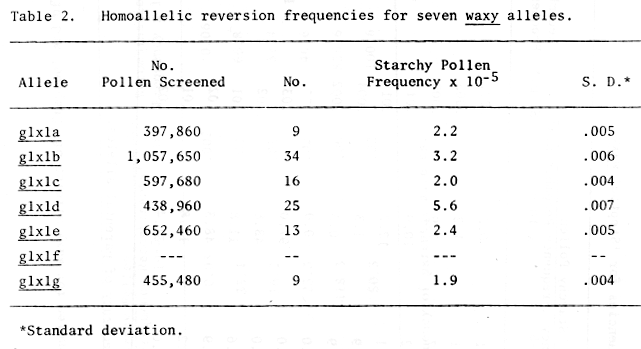
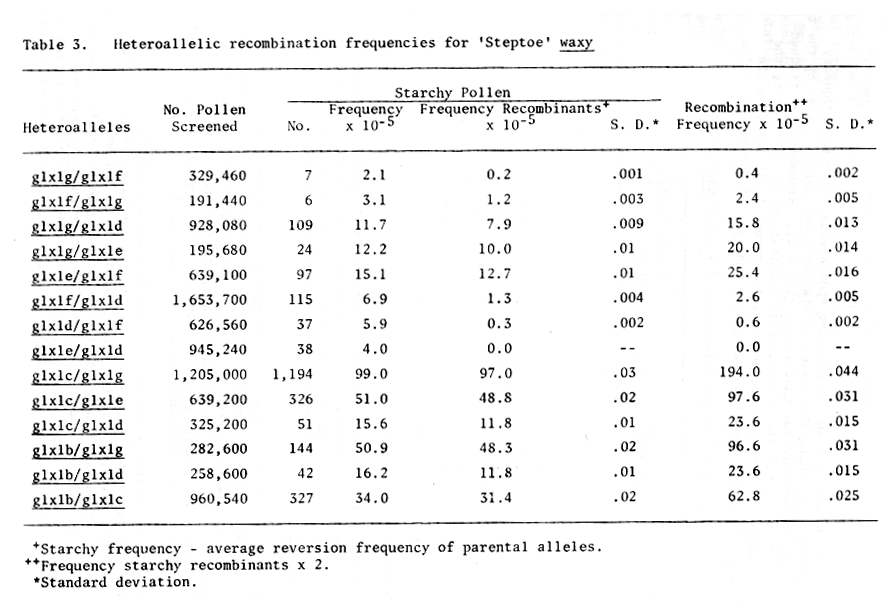
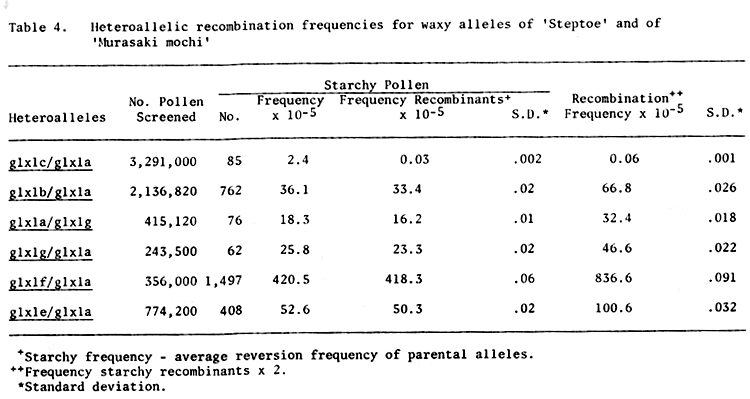
For each parental mutant allele, the spontaneous reversion frequency was calculated by dividing the number of starchy (Glx) pollen grains by the total number of pollen screened for that allele (Table 2). Recombination frequencies were determined for the heteroallelic combinations by subtracting the average spontaneous reversion frequency of the parental alleles from the starchy (Glx) frequency obtained from pollen analysis of the heteroallelic combination. This value was then multiplied by two to yield the recombination frequency for the heteroallelic combination (Tables 3 and 4). These data must be considered preliminary as they were obtained from only one set of crosses. The crosses have been made again to obtain additional recombination data.
Table 2. Homoallelic reversion frequencies for seven waxy alleles.
Table 3. Heteroallelic recombination frequencies for "steptoe" waxy
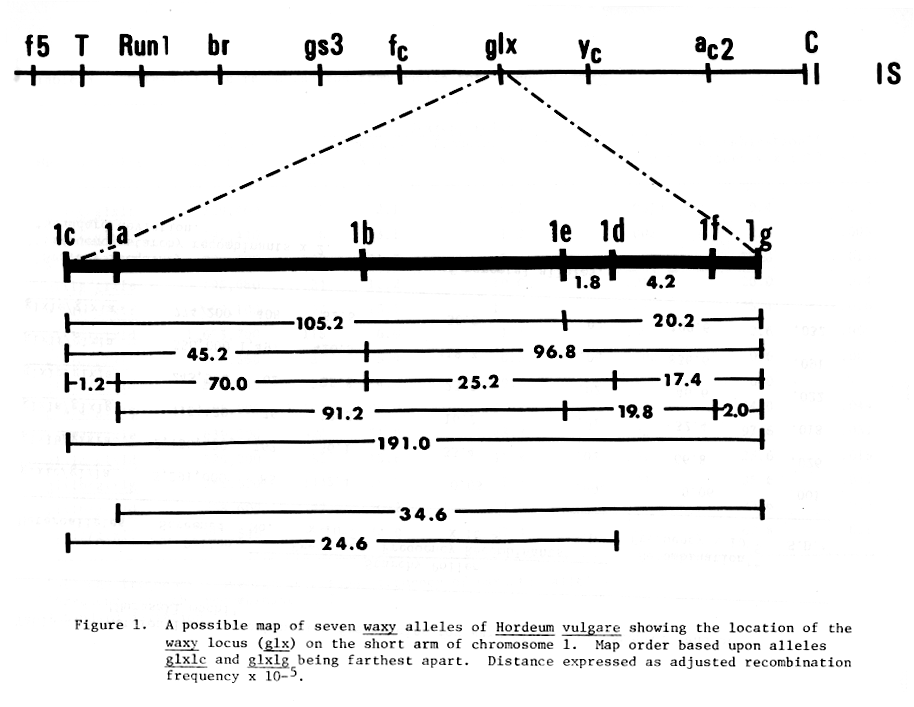
The recombination frequencies permitted construction of a preliminary map of the seven waxy alleles (Fig. 1). Several maps proved possible, and none of the maps was additive. Preliminary ordering of the alleles suggests that glxlc and glxlg are the farthest apart, and the order glxlc, glxla, glxlb, glxle, glxld, glxlf, glxlg is proposed. The mapping data do support previous reports that azide acts as a point mutagen (4,5).
The recombination values obtained can be used to estimate the genetic resolution possible in this system. The size of the waxy gene is estimated to be 900 base pairs, and if the largest (194 x 10-5) and smallest (0.4 x 10-5) recombination values are used (Table 3), the genetic resolution is 12 base pairs. This estimate may be slightly low since the waxy gene may be larger than 900 base pairs. However, even if doubled, the genetic resolution provided by this system appears to be excellent for a higher eukaryote.
To fully utilize this system for mutagen monitoring and to determine the nature of the mutations induced, tester stocks of waxy mutants are being developed that can be reverted by specific mutagens whose mode of action is known. In this way information about the mode of action of unknown environmental mutagens tested by pollen analysis can be obtained. Eventual purification and amino acid sequencing of peptide fragmcnts of the waxy gene product (UDPG starch synthetase) of wild type and mutant waxy alleles will be used to provide confirmation of tlle nature of mutations giving rise to each mutant.
References
1. Nelson, 0. E., 1968. The waxy locus in maize. II. The location of the controlling element alleles, Genetics 60:507-524.
2. Freeling, M., 1976. Intragenic recombination in maize: Pollen analysis methods and the effect of parental Adhl+ isoalleles, Genetics 83:701-717.
3. Kleinhofs, A., R. L. Warner, F. J. Muehlbauer and R. A. Nilan, 1978. Induction and selection of specific gene mutations in Hordeum and Pisum, Mutation Res., 51:29-35.
4. Kleinhofs, A., C. Sander, R. A. Nilan and C. F. Konzak, 1974. Azidc mutagenicity--mechanism and nature of mutants produced, in: Polyploidy and Induced Mutations in Plant Breeding, International Atomic Energy Agency, Vienna, pp. l95-l99.
5. Nilan, R. A., E. G. Sideris, A. Kleinhofs, C. Sander and C. F. Konzak, 1973. Azide--A potent mutagen, Mutation Res., 17:142-144.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}