II. 36 Haploids and hybrids following interspecific crosses.
N.C. Subrahmanyam, Genetics Department, Research School of Biological Sciences, The Australian National University, Canberra, A.C.T. 2601. Australia. "R"
Selective chromosomal elimination in Hordeum interspecific hybrids has gained importance as a potential tool for producing haploids (see Kasha, 1974) and to understand the underlying principles. Based on the distribution of this unique phenomenon among Hordeum interspecific crosses, a hierarchy (pecking order) of chromosome elimination was proposed (Subrahmanyam, 1977). To further examine the distribution of chromosome elimination leading to haploid formation in other combinations, crosses between different species are being made following earlier techniques (Subrahmanyam, 1977).
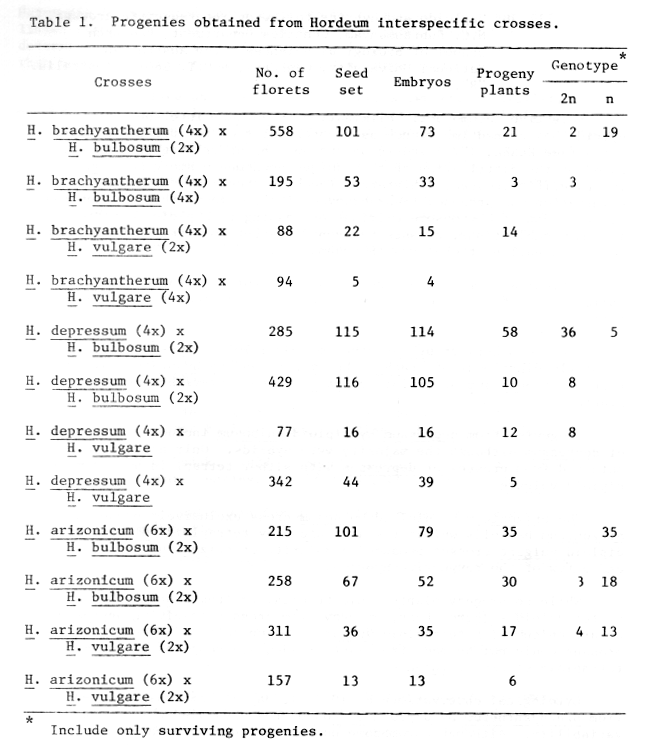
Progenies from various crosses are presented in Table 1 which indicate that five out of the twelve combinations lead to haploid formation.
Table 1. Progenies obtained from Hordeum interspecific crosses.
The cross between brachyantherum and diploid bulbosum resulted in mostly dihaploids of brachyantherum whereas brachyantherum by tetraploid bulbosum or diploid vulgare crosses invariably produced hybrids.
Progenies from depressum by diploid bulbosum included 12% haploids of depressum although the majority were hybrids. Only hybrids were obtained from crosses of depressum with either tetraploid bulbosum or diploid vulgare.
The arizonicum by diploid bulbosum cross exclusively produced arizonicum haploids whilst the arizonicum by tetraploid bulbosum or diploid vulgare crosses produced a majority of arizonicum haploids and a few of the respective hybrids.
While no progeny plants could be obtained from brachyantherum by tetraploid vulgare cross, progeny from crosses involving depressum or arizonicum with tetraploid vulgare died on potting. Reciprocal crosses could not be made from want of adequate supplies of autotetraploid stocks of vulgare.
Cytological observations on 10 to 12 day old embryos obtained from brachyantherum by diploid bulbosum cross revealed chromosome variability. Although chromosome number variants were absent in hybrid embryos from brachyantherum by diploid vulgare or tetraploid bulbosum, micronuclei were seen in some cells which indicates a certain degree of instability. Chromosome variability was also evident in one of the 6 hybrid embryos squashed from the depressum by diploid bulbosum crosses and no instability was apparent in other combinations (Table 1) involving depressum. The general pattern of variability in embryos from crosses involving arizonicum with either bulbosum or vulgare was similar to an earlier report (Islam and Sparrow, 1974). The minimum number of chromosomes in any hybrid embryo was either 14 in crosses involving brachyantherum/depressum or 21 in arizonicum crosses. This clearly suggests that chromosome elimination leads to haploid formation in the haploid-producing crosses.
These results are consistent with the generality for Hordeum that a specific ratio of the parental genomes (or chromosomes) in the zygote determines the stability or elimination of chromosomes and hence the nature of the resulting progeny. Present results, in conjunction with earlier data (Subrahmanyam, 1977), suggest differences in the "genetic strength" between different species in eliminating the bulbosum chromosomes.
Whether such differences are manifested through the rate and/or magnitude of elimination, remains to be studied.
Acknowledgements:
Thanks are extended to Dr. D.R. Dewey, Utah State University, Logan, USA and Dr. W. Lange, Wageningen, The Netherlands, for the supply of the species. The technical assistance of Miss K. Ferguson is highly appreciated.
References:
Islam, R. and D.H.B Sparrow. 1974. Chromosome elimination in interspecific hybrids involving Hordeum arizonicum (6x) x H. vulgare (2x) and H. arizonicum (6x) x H. bulbosum (2x). Barley Genet. Newsl. 4:36-37.
Kasha, K.J. (ed.) 1974. Haploids in Higher Plants - Advances and Potential.
Subrahmanyam, N.C. 1977. Haploidy from Hordeum interspecific crosses I. Polyhaploids of H. parodii and H. procerum. Theor. Appl. Genet. 49:209-217.
{kind=link}