II. 32 Trisomic Analysis of the gene f3 for chlorina 3.
Shala, Azizeh and T. Tsuchiya, Department of Agronomy, Colorado State University, Fort Collins, Colorado 80523, U.S.A.
*Partly supported by the research grant from National Science Foundation BG30493 and Colorado State University Experiment Station Project (Hatch 8).
The gene f3 for chlorina 3 mutant was first described by Immer and Henderson (1943) (Robertson et al., 1947). These authors could not find linkage of f3 with then known marker genes in any of the seven linkage groups. The analysis of allelic relationship of f3 with all other established genes for chlorina (fc, f, f2, f4, f5, f6, f7, f8, f9), and light green (lg5) proved that f3 was not allelic to any of those ten genes (Tsuchiya et al., 1973; Tsuchiya, 1974). Primary trisomic analysis of f3 was conducted with Triplo 4 (Robust) with results showing a disomic ratio (Tsuchiya et al., 1976).
In this report the results of further studies of f3 by means of primary trisomic analysis are reported. The primary trisomic stocks used in this experiment were obtained in a two-rowed cultivar Shin Ebisu 16 (SE 16) by Tsuchiya (1967). The trisomic plants were used as the female parent in crosses with homozygous f3 plants as the male parent. In order to confirm previous results on Triplo 4 - f3 relationship, the trisomic F1 plants between Triplo 4 and f3, were back crossed with f3 homozygotes as the female. The diploid F1 plants were also back crossed to f3 homozygotes as the female parent for the control. Since it has been proven that almost no pollen transmission was recorded in barley trisomics (Tsuchiya, 1967) chromosome numbers in the progenies of trisomic back crosses were not counted. The segregation ratios are shown in Table 1.
The results shown in Table 1 clearly showed that the segregation ratio was disomic in both backcrosses, confirming the previous results by Tsuchiya et al. (1976) that the gene f3 is not located in chromosome 4.
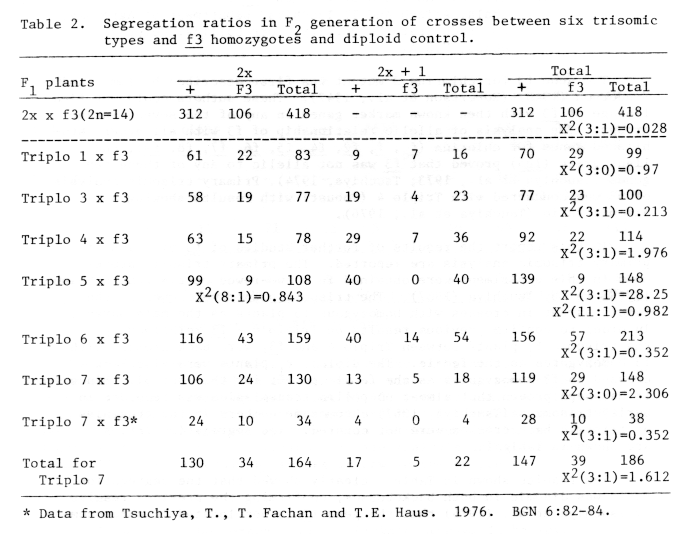
Segregation ratios were studied in F2 generation of crosses between six trisomic types and f3 homozygotes. The results are shown in Table 2 together with the segregation ratio in the diploid control.
Table 2. Segregation ratios in F2 generation of crosses between six trisomic types and f3 homozygotes and diploid control.
The results given in Table 2 indicated that the segregation ratios fit very well to the disomic ratio of 3:1 in trisomic types for chromosomes 1, 3, 4, 6, and 7, and showed that the gene f3 is not associated with any of these five chromosomes.
The segregation ratio in the F2 population of the cross Triplo 5 x f3 did not fit to 3:1 ratio but fit to 8:1 in 2x portion. Also there was no recessive homozygote in 40 plants of 2x + 1 portion (Table 2). It is, therefore, safe to conclude that the gene f3 is associated with chromosome 5.
References:
Immer, F.R. and M.T. Henderson. 1943. Linkage studies in barley. Genetics 28:419-440.
Robertson, D.W., G.A. Weibe, and R.G. Shands. 1947. A summary of linkage studies in barley: Supplement I, 1940-1946. J. Am. Soc. Agron. 39:464-473.
Tsuchiya, T. 1967. The establishment of a trisomic series in a tworowed cultivated variety of barley. Can. J. Genet. Cytol. 9:667-682.
Tsuchiya, T. 1974. Further results of allelism testing in barley. BGN 4:82-85.
Tsuchiya, T., T. Fachan and T.E. Haus. 1976. Primary trisomic analysis of lg3 (light green) and f3 (chlorina) in barley. BGN 6:82-83.
Tsuchiya, T., D.A. Jensen, and T.E. Haus. 1973. Allelism tests of some chlorina mutant genes of barley. BGN 3:71-72.
{kind=link}
{kind=link}