II. 29 Mitotic Chromosomal Aberrations in Barley Induced by Accelerated Seed Aging.
M. Murata, T. Tsuchiya, Department of Agronomy, Colorado State University, Fort Collins, Colorado 80523, and E.E. Roos, National Seed Storage Laboratory, USDA, Fort Collins, Colorado 80523.
Chromosomal aberrations are known to accumulate during seed aging, particularly under conditions of high temperature and/or seed moisture content. In this study, the effects of a severe aging treatment on seed of Hordeum vulgare cv. Himalaya (initial moisture 6.7% fresh weight basis; initial germination 100%) were investigated. Subsamples of 260 seeds per treatment were put into cans of 5.5 cm diameter x 5.5 cm height with enough distilled water to raise the seed moisture content to 18% absorbed in blotter paper and sealed in the can. The seeds were not allowed to come into direct contact with the wet blotter. After 3 days equilibration at 21 C, five sealed cans were stored at 38 C constant temperature for from 4 to 20 days. At 4-day intervals one can was removed from storage and ten seeds per can were used to determine seed moisture content by oven drying at 103 C for 24 hours. Two hundred seeds were germinated on blotter paper in petri dishes in the dark at 20 C. Beginning 20 hours after the start of imbibition, primary roots of each germinated seed were collected, their lengths measured, and the roots fixed in acetic alcohol (1:3). After 24 hours fixation, the roots were transferred into acetocarmine staining solution(0.8% acetocarmine in 45% acetic acid). After at least 3 days, squash preparations were made and chromosomal aberrations were observed only at mid-anaphase of the first mitotic division.
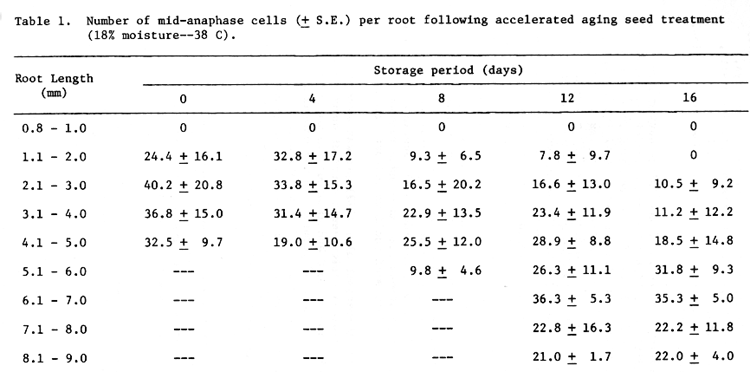
In a previous report (Murata et al., 1977), it was shown that the first mitoses in normal Himalaya barley seeds occur at about 16-18 hours after beginning of imbibition at 20 C and at about 1.5 mm in root length. In aged seeds, however, the start of germination was delayed. The 0 (control) and 3-day storage seeds started germination in less than 30 hours, whereas around 48 hours were required for initiation of germination of the seeds stored 8 days, and more than 60 hours for those stored 12 and 16 days. Therefore, the time of first mitosis and degree of cell activity were estimated on the basis of root length.
The relationship among the average number of mid-anaphases, root length, and the storage period is shown in Table 1. In the 0 and 4-day stored seeds, numerous anaphases were first observed in roots 1.1 to 2.0 mm in length and maximum numbers were noted in roots 2.1 to 3.0 mm long. With increased length of storage from 8 to 16 days, the maximum number of anaphases occurred at longer root lengths, and the number of anaphases in the 1.1 to 2.0 mm long roots were reduced. This shows that initiation of mitotic divisions in root meristems is delayed on the basis of root length as well as time from the start of imbibition. These results suggest that the aging treatment affected the synchronization of cell division.
Table 1. Number of mid-anaphase cells (+ S.E.) per root following accelerated aging seed treatment
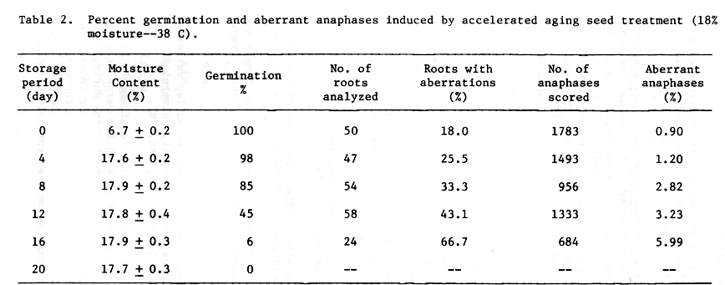
Germination decreased from 100 to 0% (Table 2) as the storage period increased from 0 to 20 days. The percentage of roots with chromosomal aberrations and aberrant anaphases increased as germination decreased, with both showing highly negative correlations (r = -0.98** and -0.97**, respectively).
These results suggest that the decrease in germination was associated with chromosome damage during artificial aging.
Roberts et al. (1967) and Abdalla and Roberts (1968) reported a similar relationship between the loss of viability and the accumulation of chromosome damage in barley, broad beans, and peas.
Aberrations observed in this experiment were almost all single-fragments (31% of aberrant anaphases observed in aged seeds) and single-bridges (46%), but the combination of fragments and bridges in the same cell was very rare (4%). This suggests that possibly the cytological damage related to loss of germination was of the chromatid-type rather than of the chromosome-type breakage. This agrees with the result in natural aged seeds of Triticum durum (Avanzi et al., 1969 and Innocenti and Avanzi, 1971). However, the chromatid-type damage in aged seeds does not agree with the chromosome-type damage reported by Abdalla and Roberts (1968) and others. In order to clarify this difference on the type of aberrations, further critical analyses are necessary.
References:
Abdalla, F.H. and E.H. Roberts. 1968. Effects of temperature, moisture, and oxygen on the induction of chromosome damage in seeds of barley, broad beans, and peas during storage. Ann. Bot. 32:119-136.
Avanzi, S., A.M. Innocenti, and A.M. Tagliasacchi. 1969. Spontaneous chromosome aberrations in relation to seed dormancy in Triticum durum Desf. Mutation Res. 7:199-203.
Innocenti, A.M. and S. Avanzi. 1971. Seed aging and chromosome breakage Triticum durum Desf. Mutation Res. 13:225-231.
Murata, M., E.E. Roos, and T. Tsuchiya. 1977. Analysis of the first mitotic divisions in germinating seeds of barley. BGN 7:81-84.
Roberts, E.H., F.H. Abdalla, and R.J. Owen. 1967. Nuclear damage and the aging of seeds with a model for seed survival curves. Symp. Soc. Exp. Biol. 21:65-100.
{kind=link}
{kind=link}