II. 24 Cytological estimate of "crossing over potentials" in primary trisomics of barley.
D.K. Khurana(1) and S. Ved Brat (2). Botany Department, Panjab University, Chandigarh, India.
Present Addresses:
1. Department of Forestry, H.P. University, Tank Road, Solan, India.
2. Department of Agronomy, Univ. Kentucky, Lexington, U.S.A.
(Note: Greek symbols in text below have been replaced with English equivalents, italicized & in quotes)
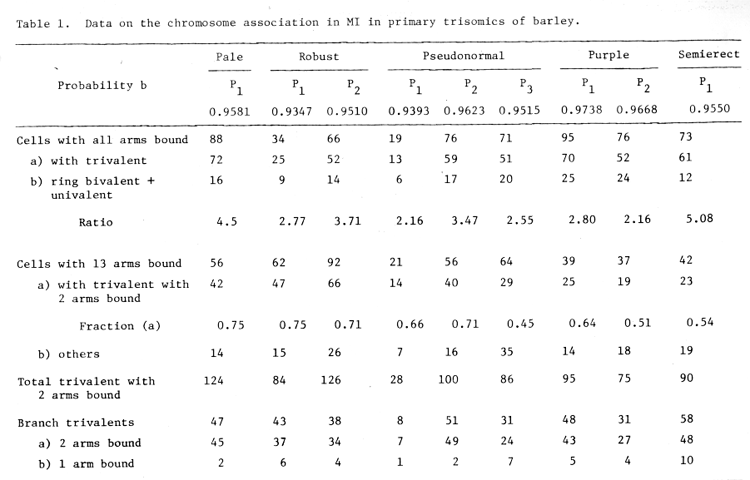
The "crossing over potentials of various chromosomes in the primary trisomics of barley was carried out according to the formula's given by Sybenga (1966a, b). The data is tabulated in Table 1. From Table 1 it is evident that for the cells with all arms bound, the ratio between those with trivalent and those without trivalent is variable and ranges between 2.16 to 5.08. As the ratio is more than 2 and variable, it shows the random pairing and not localized pairing (Sybenga, 1966b).
Table 1. Data on the chromosome association in MI in primary trisomics of barley.
This can be due to the following three reasons:
i) All the chromosome arms have equal change of pairing with on another
ii) Partner exchange takes place less frequently than in rye, therefore giving rise to high frequency of trivalents and resulting in lesser number of Y-trivalents and ring bivalents with univalents.
iii) All the 3 arms have the same tendency of pairing with one another, no preferential pairing takes place, i.e. when one arm is paired with one chromosome, the other arm not necessarily may tend to pair with the same chromosome.
Partner exchange reduces the chiasma frequency of trivalents (Sybenga, 1966b). As said earlier that the partner exchange is less frequent in barley, this is confirmed:
a) As the cell with 14 bound arms are in excess (on the whole) which is evidently the maximum number of arms to be bound. This indicates that the chromosomes have strong tendency of being bound. This is so in all cases except in Robust (Plant 1), where the excess of cells is with 13 bound arms (Table 1).
b) In the cells with 13 bound arms (Table 1) the frequency of trivalent is again more, indicating strong nature of pairing.
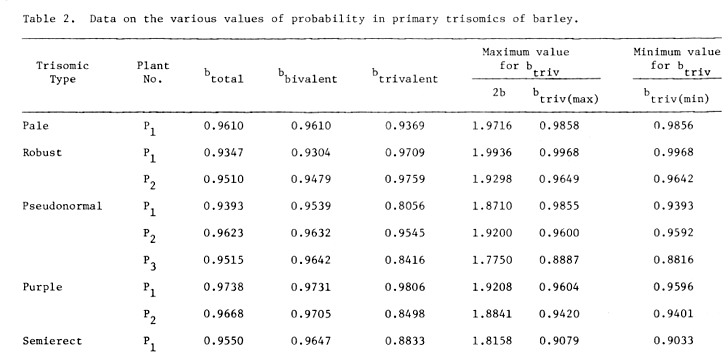
c) The proportion of arms bound in cells with a trivalent, but excluding the trivalent itself (bbiv) is slightly different that the proportion of bound arms of cell population as a whole (btotal) again indicating that the trivalents have approximately the same tendency of pairing as bivalents (see Table 2).
Table 2. Data on the various values of probability in primary trisomics of barley.
The adjusted values of btri, "mu", "nu" and maximum and minimum values for btri are given in Table 2, as per the formula of Sybenga (1966b). From the Table 2 it is evident that the maximum and minimum value of probability of pairing in Robust is similar to other trisomics, but it differs from them in having 13 bound arms in excess and having lower frequency of trivalents in 13 bound arm cells (Table 1). Therefore, it appears, that the extra chromosome in case of Robust is making the selective pairing. As the chiasma formation is under genetic control (Rees, 1955), so, perhaps the gene responsible for this is located in the Chromosome 4.
However, the values of "crossing over potentials" of both the arms of these chromosomes were not attempted due to the errors suggested by Sybenga (1966b) in the absence of telocentric trisomics for these chromosomes.
Acknowledgements:
Thanks are due to Dr. T. Tsuchiya, Colorado State University, U.S.A. for kindly making available the seeds of various trisomics.
References:
Rees, H. 1955. Heredity 9,93-116.
Sybenga, J. 1966a. Genetica 36,243-252.
----. 1966b. Genetica 36,339-350.
{kind=link}
{kind=link}