II. 7 The analysis of barley genomes. II: A molecular approach
T. Chakrabarti, C. H. Doy and N. C. Subrahmanyam, Genetics Department, Research School of Biological Sciences, The Australian National University, Canberra, A.C.T. 2601 Australia. "R"
Studies to ascertain the relationship between different species of Hordeum have so far been restricted to meiotic chromosome pairing. With the advent of new techniques in molecular biology, one can achieve more meaningful estimates of the extent of relatedness or divergence between species by subjecting DNA(s) to physicochemical analysis. As a first step towards this goal, a set of three related diploid species H. vulgare, H. agriocrithon and H. spontaneum, and another set of three species (sharing a genome in common and forming a polyploid series, Rajhathy et al. 1963) H. californicum (2x), H. jubatum (4x) and H. arizonicum (6x) were chosen.
DNA was isolated according to Marmur (1963) from either shoots or leaves of glasshouse grown plants and routine checks were made for bacteria in the respective extracts. The level and type of bacteria in all the species studied were similar to that from H. vulgare following AgNO3 treatment (Chakrabarti et al., 1978).
The DNA of all these species form a single band in neutral caesium chloride (CsCl) density gradients and have a buoyant density as that of H. vulgare (1.701 + 0,001 gem-3). This suggests that the repeated sequences in these species remain undetected within the mainband. However, certain classes of repeated sequences are known to be shifted and form separate bands (satellites) only when bound with antibiotics (actinomycin D or netropsin) or heavy metals, silver (Ag+) or mercury (Hg++) and run in CsCl or CsS04 gradients respectively.
In the present experiments we chose Ag+/CsS04 gradients to study the satellite pattern and to further purify satellites. When DNA from H. vulgare L. cv. Clipper was run in an analytical Ag+/CsS04 gradient at an rf (Ag+/DNA phosphate ratio) of 0.2, two satellites were found only on the heavy side of the mainband as reported by Ranjekar et al. (1976). At least two additional peaks appear on the light side of the mainband when the DNA is run at rf 0.3. Since the associated bacterial DNA is not displaced in Ag+/CsS04 gradients, it is highly unlikely that the bacterial DNA contributes to the present satellites. Following fractionation on a preparative Ag+/CsS04 gradient and subsequent analytical ultracentrifugation in neutral CsCl, the buoyant densities of individual satellites were obtained. The provisional values are: 1.724, 1.701 and 1.700 from the heavy side and from the light side, one at 1.702 with another which may have three components, 1.692, 1.701 and 1.706 gcm-3. These satellites together comprise at least 5% of the total DNA.
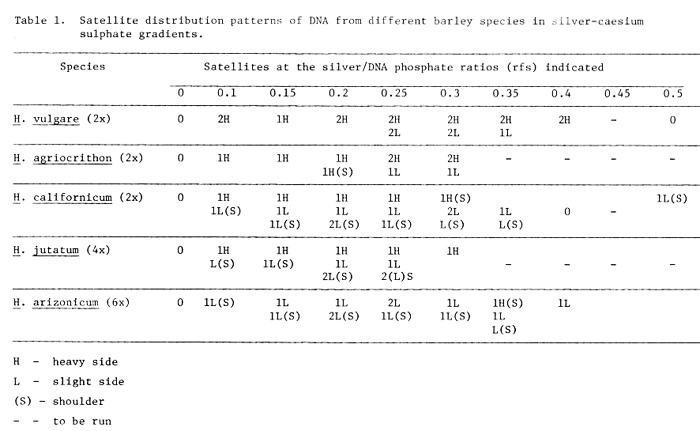
The satellite distribution patterns in other species over a range of rfs are presented in Table 1. These results clearly indicate differences in the gross pattern between species, the most obvious difference being that each of the related diploid species have two heavy peaks, whereas only one is present in any of the species in the polyploid series. Furthermore, a comparison of the number of satellites resolved at different rfs clearly suggests that different rfs are necessary in order to achieve the best separation of the satellites in different species.
Experiments have commenced to determine the buoyant densities of different satellites for each of these species, their chromosomal locations, reassociation patterns, their relationships and sequence diversity in terms of restriction endonuclease analysis. In-situ hybridization of "species specific" satellites might become useful to identify chromosomes (genomes) belonging to different species.
Acknowledgements
We wish to thank Dr. D. R. Dewey of Utah State University, Logan, Utah, U.S.A.; Dr. R. A. Finch, PBI, Cambridge and Dr. D. H. B. Sparrow for supplying seeds of different species used in the present study. Assistance of Mr. M. Perri in running the analytical ultracentrifuge is highly appreciated.
References:
Chakrabarti, T., C. H. Doy and N. C. Subrahmanyam, 1978. The analysis of barley genomes I. The problem that the DNA of bacteria may contribute to the DNA in extracts of barley tissues derived from germinated seeds. Barley Genetics Newsletter 8:
Rajhathy, T., J. W. Morrison and S. Symko, 1963. Interspecific and intergeneric hybrids in Hordeum. Barley Genetics J.: 195-213.
Ranjekar, P. K., D. Pallotta and J. G. Lafontaine, 1976. Analysis of the genome of plants II: Characterization of repetitive DNA in barley (Hordeum vulgare) and wheat (Triticum aestivum). Biochim. Biophys. Acta. 425: 30-40.
{kind=link}