IV. 6 Coordinator's report: Chromosome 6.
K.J. Kasha. Crop Science Dept., University of Guelph, Guelph, Ontario, Canada N1G 2Wu
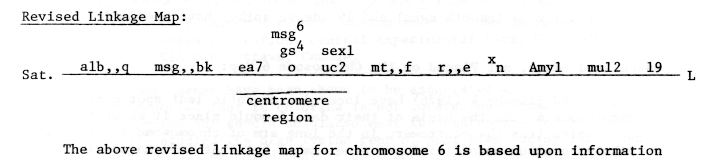
The above revised linkage map for chromosome 6 is based upon information from previous issues of BGN plus two articles in the current issue Vol. 8, namely those of Konishi and of Kasha, Falk and Tsai. The orientation of the linkage map is based on the chromosomal interchange linkage data of Ramage and Paluska (BGN5:49-51) which located msg,,bk in the short arm, about 6 c.o. units from the centromere. While gene distances have not been included at this stage, the illustrated map indicates that there are about six genes located close to the centromere region of chromosome 6, perhaps within 4 c.o. units of each other. Their gene order is not clear except that ea7 is probably to the left and sex1 and uc2 to the right of o. The gene order of Amyl and mul2 in the long arm is tentative and based upon their recombination values with o, obtained in separate studies.
The major problem is the wide variation in recombination values from cross to cross and from independent studies by various researchers. At least a two-fold difference is not uncommon, even within crosses handled under the same environments at the same location (see 3 point data of Kasha et al., BGN8). The data of Konishi (BGN8) show quite large recombination values between genes studied when compared with previous reports. Thus, I suspect the location of Amyl and mul2 between xn and l9 is reasonable.
There are some major changes in the linkage map of chromosome 6 when compared to previous reports. The long arm of 6 has been extensively extended with the data of Konishi (BGN8) indicating l9 may be 70 c.o units from o and the centromere. This may now allow us to explore and locate genes in the long arm which up to now could not be detected by gene linkages. The mul2 gene has been switched from the short arm to the long arm of 6. Kasha et al. (BGN8) found independence between ea7 and mul2 and in 3-point linkage data, mul2 was on the opposite side of o from msg,,bk. The linkage value (24.2 + 6.4) between o and mul2 is consistent with that reported by Walker et al. (Can. J. Genet. Cytol 5: 200-219, 1963). The 3-point data (Kasha et al. BGN8) failed to provide a consistent gene order for o and gs4 while sexl is closer to o than previously listed in BGN7.
Genes added to the Revised Map:
a) The albino seedlings, alb,,q (previously listed as alb,,f) is placed distal to msg,,bk in the short arm on the basis of 3-point data with msg,,bk and o (Kasha et al., BGN8). The population size was small and further data are required, although the value of 11.7 of alb,,q to msg,,bk is close to the 13.2% obtained by Rahman and Eslick (BGN6:53-58).
b) Eslick, Ramage and Clark (BGN4:11-15) placed msg6 about 1 c.o. unit from o so it has been placed in the centromere region.
c) On the basis of the data of Konishi (BGN8) relative to previous estimates of recombination between o and xn (range 6 to 18%), the genes mt,,f (mottled), r,,e (smooth awns) and l9 (dense spike) have been added to the map.
Associated Genes, not Placed on the Chromosome 6 Map:
a) Hauser and Fischbeck (1976) have located a necrotic leaf spot gene on chromosome 6. On the basis of their data I would place it about 6 to 12 c.o. units from the centromere in the long arm of chromosome 6. It will likely be given an official nec gene symbol but that information is not yet available. Their 28.3% recombination of nec with o is likely a misprint since they did not recover double recessives in the F2. Using maximum likelihood formulae, my calculations are less than 1%, or 13% if you add one to the double recessive class.
b) The piebald or pale green seedling gene (Kasha and Tsai BGN7:43) is located about 7 c.o. units from o in the long arm of chromosome 6. This will place it somewhere from mt,,f to xn. It is not listed on the map because a gene symbol has not been requested and it should be allele tested with mt,,f.
c) alb,,t (albino seedings) has been associated by trisomic analysis (Clark and Ramage, BGN6:8).
d) ert e(dense spike) was associated with chromosome 6 (see Nilan, 1963, Cytol. and Genetics of Barley) but was found to be independent of msg,,bk and msg6 (Eslick, Ramage and Clark, BGN4:11-15). It could be in the long arm and should be allele tested with l9.
Two other genes, cu (curly awns) and rb (ribbon grass) were associated with chromosome 6 on distant linkages using only F2 data. So far these genes have not been located on other chromosomes and perhaps they are distal in the long arm in spite of the failure to find linkage with o (Kasha et al. BGN8). These genes and ert e need to be checked against trisomics for 6 and should be crossed with l9, mul2 and Amy1.
Odds and Ends: Telotrisomics for chromosome 6 would also be valuable to sort out the arms of the many genes near the centromere, but these are not available.
Various multiple marker gene stocks are now available upon request. Workers report difficulty in classifying mul2 and ea7 in some crosses and others find them easy to classify. The penetrance of r,,e may be low in some instances. The shrunken endosperm gene sex1 looked good in the field at Guelph last year and may be a most useful gene in that F3 classification can be made on seed from F2 plants, thus saving a generation. If you are sending information on chromosome 6 for inclusion in BGN, please also send me a copy so that the Coordinator's Report will not be a year behind in assembling the information.
Reference:
Hauser, J. and G. Fischbeck. 1976. Z. Pflanzenzuchtg.,77: 269-280.
{kind=link}