III. 1 A proposal for sex-marked interchanges to assign genes to barley chromosomes.
D.E. Falk and K.J. Kasha, Crop Science Dept., University of Guelph, Guelph, Ontario, Canada N1G 2W1.
The proposed method combines shrunken endosperm genes, which express xenia (sex), with selected chromosomal interchanges to provide a tester set for locating genes as to chromosome in barley. Genes which express xenia, such as sex1 and sex3, are particularly useful in linkage studies in that the homozygous recessive F2 seeds can be identified and separated from the dominant phenotypes before planting the F2 population. By planting these recessive F2 separately, one can rapidly check for linkage with other genes by observing their segregation in the two sub-populations. Furthermore, the F3 classification for sex can be made on seed from the F2 plants so that more accurate and reliable linkage estimates may be obtained without growing the F3 generation.
Heterozygous chromosomal interchanges in barley show greatly reduced recombination values in the interstitial segments (the regions between the interchange breakpoints and the respective centromeres). The gene for sex1 is located very close to the centromere on chromosome 6 (see Foster, BGN 7: 28-29 and Chromosome 6 Co-ordinators Report in BGN 8) while sex3 is apparently close to the centromere on chromosome 7 (see Ullrich and Eslick, BGN 7: 66-73; Karlsson BGN7: 40-42). Therefore, both genes will show close linkage with any interchanges involving their respective chromosomes and pseudolinkage with any genes in the interstitial segment or near the breakpoints or centromeres of the other chromosome involved in the interchange.
Our proposal involves the selection of a set of interchanges in barley which would mark the various chromosomes, preferably each arm, and incorporating the sex genes into them. The selected interchanges should have breakpoints in the middle or sub-terminal regions of the chromosome arms to permit detection of linkage with genes in most regions of the chromosomes (Fig. 1).
Some comparisons of the proposed system with the often used set of primary trisomics follow:
1. No cytology and fewer F1 seed are required with our system whereas trisomic F1 plants must be identified and selected in the trisomic system.
2. Homozygous interchange stocks usually exhibit normal plant vigor and can be used as male or female parents in crosses. The former is not true of trisomics and the latter is useful for locating genes such as the male steriles.
3. Using our system, genes should segregate 3:l in all F2 populations and more accurate linkage information can be obtained using smaller populations when compared to trisomics.
4. While our system would mark most chromosome regions (Fig. l), the probability of association with a specific chromosome is greater using trisomics.
Other features or advantages of our proposed system are:
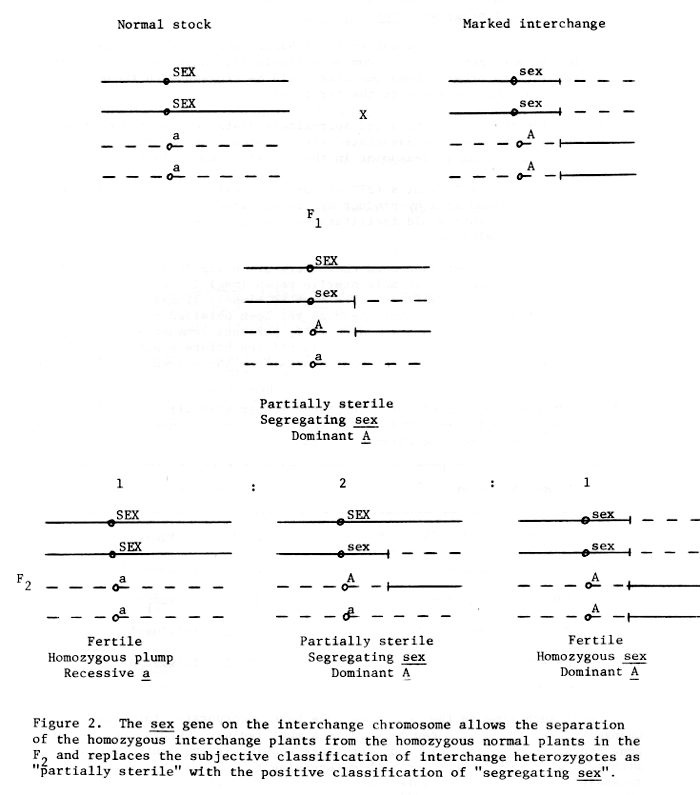
1. Since linkage would be detected by segregation relative to sex, the sometimes difficult classification of partial sterility due to the heterozygous interchange is not required (Fig. 2).
2. Due to the close linkage between sex and the interchanges, one could separate most normal and interchange homozygotes in the F2 if one wished to use interchanges in linkage calculations.
3. Regular maximum likelihood equations (3:6:3:1:2:1 ratio) may be used to calculate linkage with sex.
4. Crosses to a set of 5 sex-marked interchanges should be sufficient to locate most genes as to chromosome (Table 1). A second series with breakpoints in other chromosome arms could be crossed with those genes failing to show linkage with the first set.
5. To provide the arm location and approximate distance from the centromere of genes that have been associated with a chromosome, a cross with an interchange having a breakpoint in the opposite arm could be made.
6. Balanced Tertiary Trisomics (BTT's) for the previously unlocated gene could be obtained as a by-product of crosses involving the sex-marked interchanges. This would facilitate further linkage studies for lethal genes such as albinos.
When genes for other than seed characters are being located, the tester set could also contain genetic male sterile genes (msg) to facilitate crossing onto the tester set (Table 1). The msg6 is closely linked to sex1 on chromosome 6 while no recombinants have as yet been obtained between msg19 and sex3 on chromosome 7 (Eslick and Hockett, personal communication). Since sex homozygous recessives could be identified before planting the F2 seed, it would not be serious that most plants from these seeds would be male sterile.
Until such time as the sex genes have been incorporated into the interchange stocks, the system can be used in a modified way. Cross the unlocated gene stocks to sex1 and sex3 and, for those showing independent segregation with sex1 and sex3, select the double recessive F2. Cross these double recessives with the set of interchange stocks. When linked by the interchange, the unlocated gene will show pseudolinkage with sex in the coupling phase, thus providing the chromosomal association of the unknown locus.
{kind=link}
{kind=link}
{kind=link}