II. 1 Male sterile mutants of barley. VI. Tests of restoration by the cytoplasm of Hordeum bulbosum, and reports of new mutants.
H. Ahokas, Department of Genetics, University of Helsinki, P. Rautatiekatu 13, 00100 Helsinki 10, Finland. "R"
Many barley breeders have produced Hordeum bulbosum alloplasmic barleys. This alien cytoplasm influences some quantitative and qualitative traits, such as plant height, straw stiffness, tillering capacity, growth rhythm, senescence, heading and maturity date, and yield. The alloplasmic barley behaves in some respects like a mutant sensitive to cool temperatures (FUKUYAMA and TAKAHASHI 1976). Taking advantage of male sterile genes, the search for superior combinations of barley genotypes and bulbosum cytoplasm was suggested by WIEBE (1976).
Several major genes of barley penetrate in bulbosum cytoplasm. However, it was of some academic interest to test the penetrance of male sterile genes (msg s) of barley for possible cytoplasmic restoration. By using embryo culture in vitro, I have prepared 20 bulbosum alloplasmic barley lines from cv. Karri carrying the cytoplasm of 9 different tetraploid accessions. One of these lines was prepared from a male sterile plant of H. bulbosum. This sterility was not transmitted to the alloplasmic derivative, HA-Hb-20-Karri. HA-Hb-l-Karri was used to test the segregation of male steriles in F2 families or generations (Table 1). No restoration was found. Eventual partial restoration of fertility was not studied.
Table 1. Segregation of male steriles in the cytoplasm of H. bulbosum (HA-Hb-1-Karri).
The mutants msg,,dm (P-11) and msg,,dn (H-31) are allelic (Table 2). The difference in their phenotype (see AHOKAS 1976) may be caused by the different genetic background. This locus is evidently not the same as that of the mutants msg6, msg8,or msg16 (Table 2) which also produce inaperturate pollen grains (AHOKAS 1976). Two other cases of allelism are reported in Table 2.
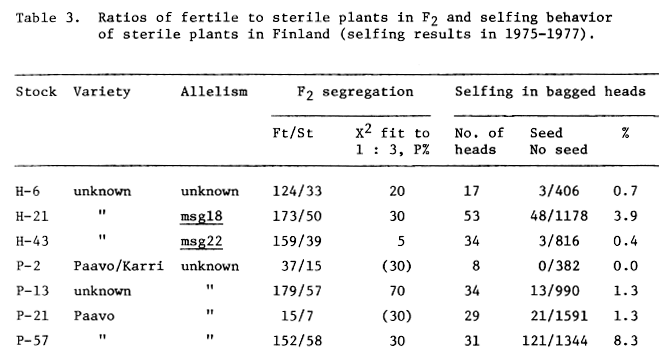
Most of the new mutants are partial male sterile (Table 3). The partial male steriles can be most usable mutants for establishing composite cross bulks, since they permit crossing and selfing.
The male sterile in P-57 shows an undescribed form of partial male sterility. The anthers are equal to or a little smaller than those of a fertile sib, but incapable to dehisce or the dehiscing is delayed. The artificial opening of the anthers with forceps increases selfing significantly to the mean level of 27.4%. Probably similar undehiscence was induced with DPX 3778 in some other cereals (JOHNSON and BROWN 1976).
Due to the negative results on cytoplasmic restoration by bulbosum cytoplasm and the unfavourable effects associated with this cytoplasm further tests are discontinued. Instead, the cytoplasmic male sterile (msm1) and its restorer of fertility (Rfm) which I have recently derived (AHOKAS 1978a and b) are expected to be beneficial to hybrid barley production.
Acknowledgements:
The work was done under the auspices of the Academy of Finland.
References:
AHOKAS, H. 1976. Male sterile mutants of barley. III. Additional inaperturate mutants. BGN 6:4 - 6.
AHOKAS, H. 1978a. Cytoplasmic male sterility in barley. Genetics (submitted).
AHOKAS, H. 1978b. Cytoplasmic male sterility in barley. I. Anther and pollen characteristics of msm1, restored and partially restored msm1. Z. Pflanzenzucht. (in press).
FUKUYAMA, T. & TAKAHASHI, R. 1976. A study of the interspecific hybrid, Hordeum bulbosum (4X) x H. vulgare (4X), with special reference to dihaploid frequency. Barley Genetics III:351-360.
JOHNSON, R. R. & BROWN, C. M. 1976. Chemical control of pollination in wheat and oats. Crop Sci. 16:584-587.
WIEBE, G. A. 1976. The challenge facing barley breeders today. Barley Genetics III:l-lO.
{kind=link}
{kind=link}
{kind=link}