II. 44 Chromosome location evidence for Risø induced high lysine shrunken endosperm mutants of barley. (1)
S.E. Ullrich and R.F. Eslick. Department of Plant and Soil Science, Montana State University, Bozeman, Montana 59717 "R".
(1) Supported in part by USA AID/ta-C-1094.
Bomi, Risø mutants 8, 13 and 1508 and Carlsberg II mutants 29, 56 and 86 display a shrunken endosperm phenotype that expresses xenia in our environments, and is associated with a high lysine trait. Doll et al. (1974) described these mutants as being high in lysine content and low in kernel weight. Subsequently, Doll (1976) demonstrated for Risø 8 and 13 that the shrunken endosperm and high lysine traits are inherited together as single recessive genes. Karlsson (1977) and Ullrich and Eslick (1977) have shown the same to be true of Risø 1508, and Ullrich and Eslick (1977) suggested the shrunken endosperm xenia trait of Risø 1508 be designated sex3c.
Risø mutants 8, 13, 1508, 29, 56, and 86 were crossed with a series of Betzes primary trisomics to assign the shrunken endosperm xenia expressing trait of these mutants to chromosomes (see Tsuchiya, 1964). It should be noted at the outset that the three F2 endosperm genotypes (Sex Sex, Sex sex, sex sex) were sometimes difficult to differentiate in crossed progeny for some mutants under some conditions. The degree of shriveling of the kernels varied somewhat with the environment. When difficulty was experienced observed, 3:1 and/or 1:2:1 ratios were tested to try to sort out discrepancies.
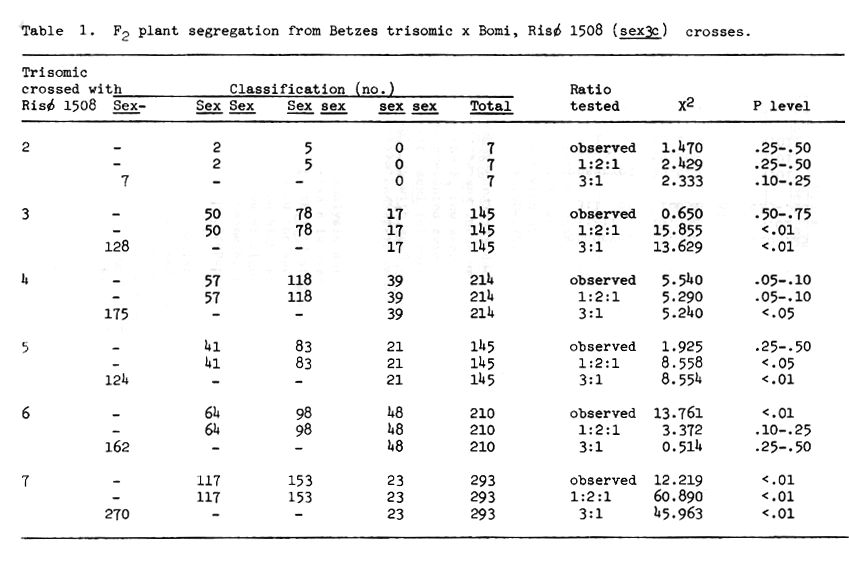
Considering the data of Table 1 in total, sex3c of Risø 1508 would be assigned to chromosome 7. This conclusion is well supported by linkage data presented by Karlsson (1977) and Ullrich and Eslick (1977).
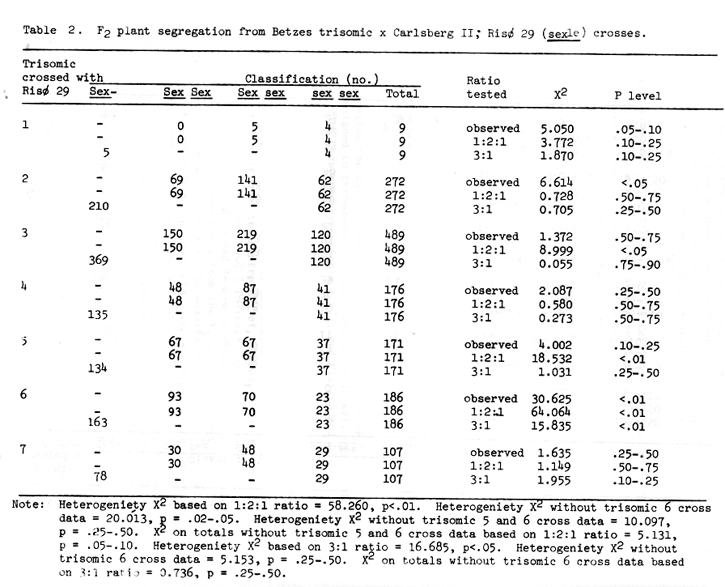
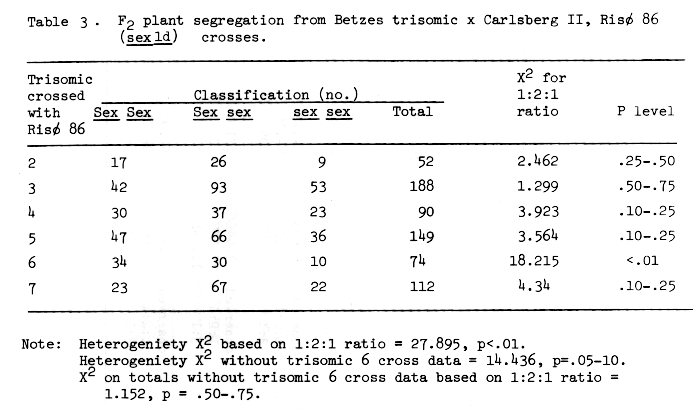
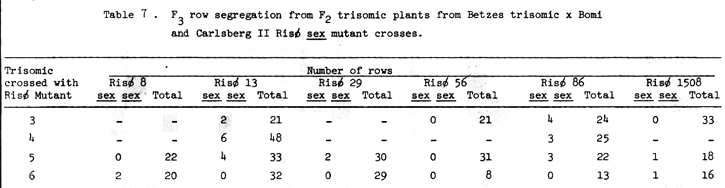
Ullrich and Eslich (1978) presented evidence that Risø 29 and 86 are allelic to each other and with Campana sex1a. Jarvi and Eslick (1975) assigned sex1 to chromosome 6, and Eslick and Reis (1976) further positioned it near the centromere of chromosome 6. The trisomic analysis data of Tables 2 and 3 indicate that the shrunken endosperm loci of both Risø 29 and 86 are on chromosome 6. Additional support for this conclusion comes from the F3 row data from F2 trisomic plants presented in Table 7. No homozygous recessive (sex sex) F3 rows are expected from F2 plants that are trisomic for the chromosome on which the gene in question is located. The specific gene action of the shrunken endosperm or high lysine characters of Risø 29 and 86 has not been reported. Since the sex traits are allelic with Compana sex1, monofactorial recessive gene action might be assumed. If the segregation of shrunken endosperm presented in Tables 2 and 3 (minus trisomic 6 cross data) are considered then single recessive gene action is concluded for the sex traits of Risø 29 and 86. Considering all the data availabe, we have proposed that the shrunken endosperm zenia trait of Risø 86 be designated sex1d and that of Risø 29 sex1e (Scheuring and Ramage, 1976, assigned sex2b, a zygotic lethal mutant, in effect, because it is available in the sex2 sex2 condition).
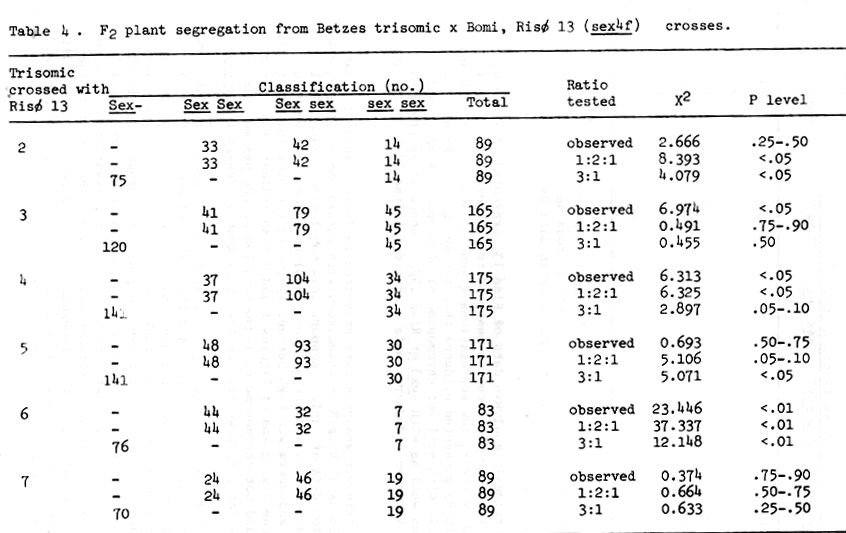
The trisomic analysis data of Risø 13, presented in Table 4, indicate that the sex locus be assigned to chromosome 6. This is supported by the F3 row segregations given in Table 7. Ullrich and Eslick (1978) presented evidence that the sex gene of Risø 13 is non-allelic with sex1 (of chromosome 6) of Compana, Risø 29 and Risø 86 as well as with sex3 of Risø 1508. Hence it is proposed that the shrunken endosperm xenia character of Risø 13 be designated sex4f.
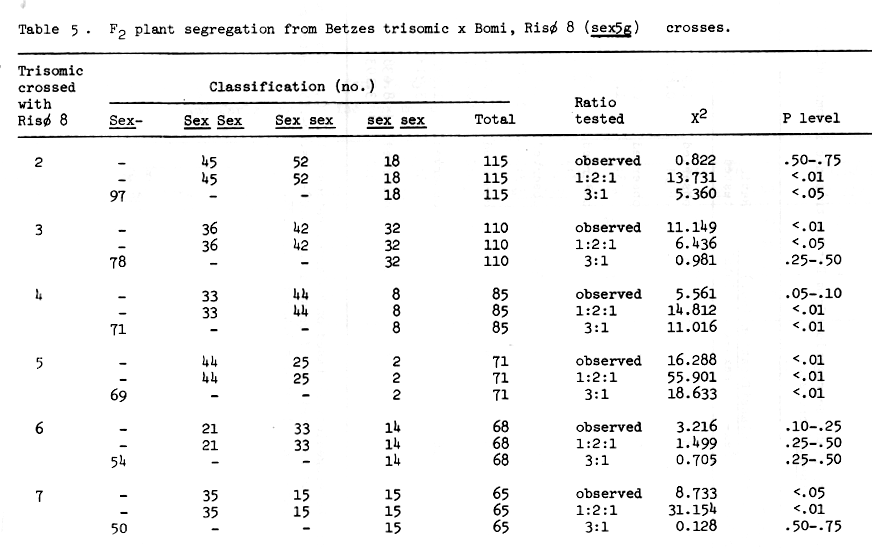
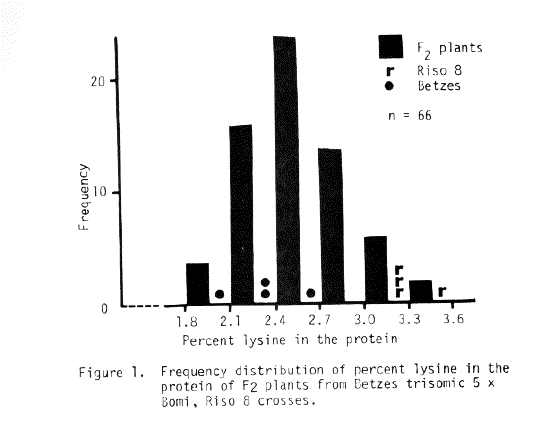
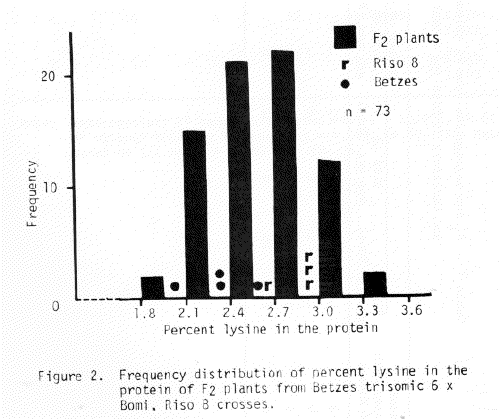
The trisomic analysis data presented in Table 5 indicate that the sex locus of Risø 8 is associated with chromosome 5. The F3 row segregation data of Table 7 support this conclusion. Since Doll (1976) has established the apparent absolute association between shrunken endosperm and high percent lysine in the protein, the lysine data in the histograms of Figures 1 and 2 also indicate that chromosome 5, and not chromosome 6, is the location of the sex mutants described to date, then we propose that this gene be designated sex5g.
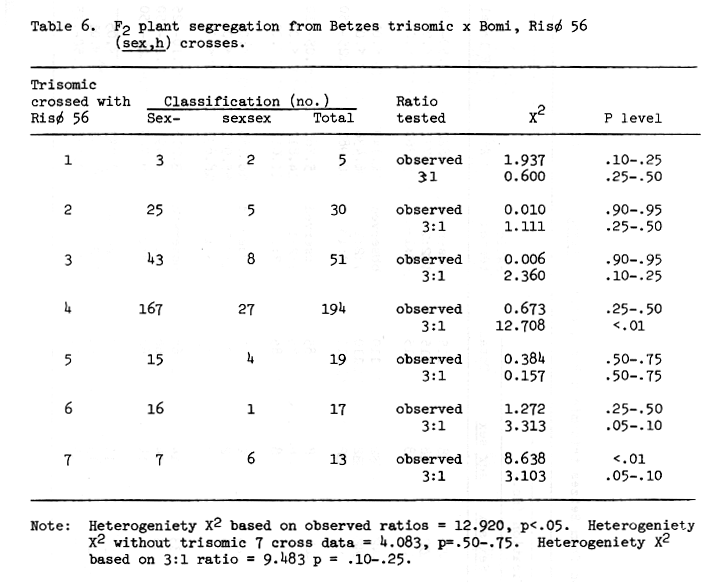
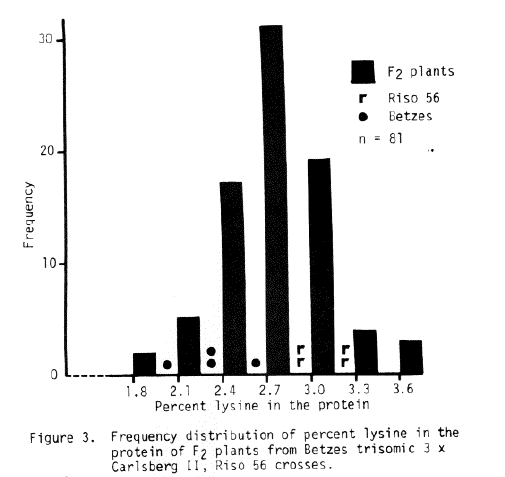
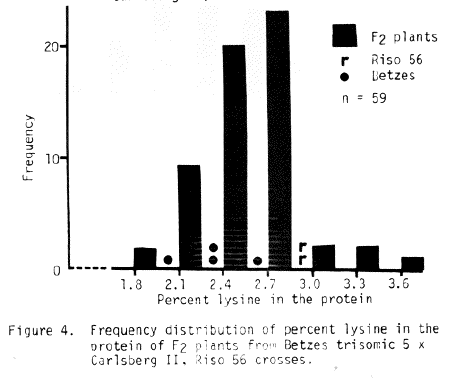
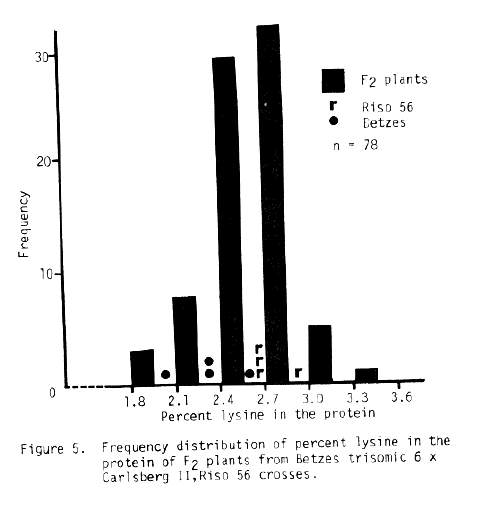
The trisomic data available for Risø 56 is sketchy at best. The sex trait is difficult to read and the number of progeny from most crosses is small. Some conclusions may be drawn, if one assumes single recessive gene action and a shrunken endosperm - high lysine association. The best evidence from Table 6 indicates chromosome 4 is the location of the sex trait of Risø 56. The lysine data presented in Figures 3, 4, and 5 indicate chromosome 5 as the associated chromosome while probably eliminating chromosomes 3 and 6. The data for Risø 56 in Table 7 perhaps supports chromosome 5 as the gene location. Additional linkage investigation is certainly indicated with perhaps an emphasis on work with chromosomes 4 and 5.
Additional linkage study is also indicated for the other sex mutants involved in this study particularly Risø 8 and 13.
Table 1. F2 plant segregation from Betzes trisomic x Bomi, Risø 1508 (sex3c) crosses.
Table 2. F2 plant segregation from Betzes trisomic x Carlsberg II; Risø 29 (sex1e) crosses.
Table 3 . F2 plant segregation from Betzes trisomic x Carlsberg II, Risø 86 (sex1d) crosses.
Table 4 . F2 plant segregation from Betzes trisomic x Bomi, Risø 13 (sex4f) crosses.
Table 5 . F2 plant segregation from Betzes trisomic x Bomi, Risø 8 (sex5g) crosses.
Table 6. F2 plant segregation from Betzes trisomic x Bomi, Risø 56 (sex,h) crosses.
References:
Doll, H. 1976. Genetic studies of high lysine barley mutants. pp. 542-546. In H. Gaul (ed.) Barley Genetics III. Proc. 3rd Intl. Barley Genet. Symp. Verlog Karl Thiemig, Munich.
Doll, H., B. Køie, and B.O. Eggum. 1974. Induced high lysine mutants in barley. Radiation Bot. 14:73-80.
Eslick, R.F. and M.N. Reis. 1976. Positioning sex1 on chromosome 6. Barley Geneti. Newsl. 6:21-22.
Jarvi, A.J. and R.F. Eslick. 1975. Shrunken endosperm mutants in barley. Crop Sci. 15:363-366.
Karlsson, K.E. 1977. Linkage studies in a gene for high lysine content in Risø barley mutant 1508. Barley Genet. Newsl. 7:40-43.
Scheuring, J.F. and R.T. Ramage. 1976. A tertiary trisomic balanced for both mgs2 and sex2. Barley Genet. Newsl. 6:60-62.
Tsuchiya, T. 1964. Chromosome aberrations and their use in genetics and breeding in barley - trisomics and anueploids. pp. 116-150. In Barley Genetics I. Proc. 1st Intl. Barley Genet. Symp. Centre for Agric. Publ. and Doc., Wagneingen, Netherlands.
Ullrich, S.E. and R.F. Eslick. 1977. Inheritance of the shrunken endosperm character, sex3c of Bomi, Risø mutant 1508 and its association with lysine content. Barley Genet. Newsl. 7:66-73
Ullrich, S.E. and R.F. Eslick. 1978. Allelism evidence for barley high lysine, shrunken endosperm xenia (sex) mutants. Barley Genet. Newsl. 8:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}