II. 40 Telotrisomic analysis of four mutant genes in barley.*
T. Tsuchiya and Laura B. Hall, Department of Agronomy, Colorado State University, Fort Collins, Colorado 80523, U.S.A.
*Partly supported by the research grant from National Science Foundation GB 30493 and Colorado State University Experiment Station Project (Hatch 8).
Four genes used in this experiment are as follows:
BGS f2 chlorina-2 recessive
28-3398 B9-11
BGS lg3 light green 3, recessive
B15-3
BGS __ short awn - incomplete dominance
T _
BGS lk5 short awn, recessive
T _
Analysis of f2: The gene f2 was studied by means of primary trisomic analysis (Tsuchiya and Robertson, 1971) and telotrisomic analysis (Singh and Tsuchiya, 1974; Tsuchiya, 1972; Tsuchiya and Singh, 1973) with the results that the gene is located in the long arm of chromosome 3. However, the occurrence of three recessive homozygotes in the trisomic portion (60 plants) of the F2 population suggested the possiblity that the gene f2 could be in the other arm of chromosome 3. Further tests have been conducted to obtain the definite association of f2 with one arm of chromosome 3.
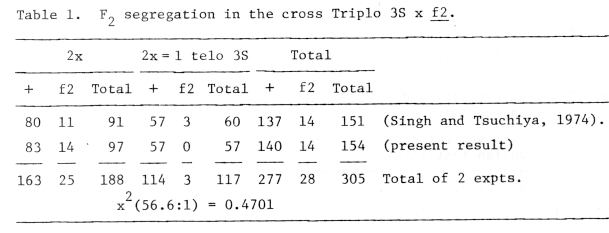
The results were shown in Table 1. This data contradicts the previous interpretation that the gene f2 is associated with the long arm of chromosome 3. The combined data (Table 1) suggest that the gene f2 is located in the distal part of the short arm of chromosome 3 (3S), although additional information is needed to reach final conclusion concerning the definite localization of this gene.
Table 1. F2 segregation in the cross Triplo 3S x f2.
Analysis of lg3: The gene lg3 has been reported to be located in chromosome 4 with the following linkage data:
Kk - Lg31g3 10.5%
Hanson, 1952, Genetics 37:90-100
Between Kk and Bl bl Hanson and Kramer 1949, Genetics
34:687-700
The primary trisomic analysis confirmed the association of lg3 with chromosome 4 (Tsuchiya et al., 1976). The gene lg3 was studied in this experiment by means of telotrisomic analysis with Triplo 4S. The results are shown in Table 2.
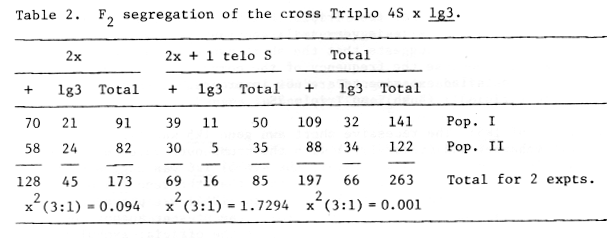
Table 2. F2 segregation of the cross Triplo 4S x lg3
The segregation ratios in 2 x portion, 2x + 1 portion and combined 2x and 2x +1, fit 3:1 very well and an appreciable number of recessive homozygotes (16 of 85 plants) were found in the 2x + 1 portion. These results clearly indicated that the gene lg3 is not associated with the telocentric chromosome used in this experiment but located in the opposite arm (4L) from f9-K arm.
Analysis of KM 200: An incompletely dominant short-awned mutant was obtained by irradiation of a two-rowed cultivar, Bakata No. 2, and named KM 200 (Tsuchiya, 1962). The homozygous mutant showed short-awn, and the heterozygotes showed medium long awn compared to the long awn in the original variety.
The mutant gene has been studied by means of primary and telosomic trisomic analysis against Triplo 1S (Tsuchiya and Haines, 1975), Triplo 3, 4, 5, 6, and 7 (Tsuchiya and Singh, 1972). The results showed that the gene is not associated with any of the chromsomes 1S, 3, 4, 5, 6 and 7 (Tsuchiya and Haines, 1975).
Telotrisomic analysis has been conducted with two telotrisomic lines, Triplo 1L and Triplo 2L. The results are shown in Table 3.
Table 3. F2 segregtaion of the crosses Triplo 1L x KM 200 and Triplo 2L x KM 200.
It is obvious that the gene is not associated with Telo 2L, since an appreciable number of recessive homozygotes were found in the trisomic portion and the segregation ratios for 2x (x2 = 0.207), 2x+1 (s2 = 0.666) and the combined 2x and 2x+1 portion (s2 = 0) fit to 3:1.
The results obtained from Triplo 1L x KM 200 were not conclusive since the population size is very small and many seed were not germinated. However, it suggests that the short awn gene may be associated with Telo 1L, because the frequency of the homozygotes was extremely low. More detailed experiments are now in progress with the primary trisomic (Triplo 1) (Bush) and Triplo 2S.
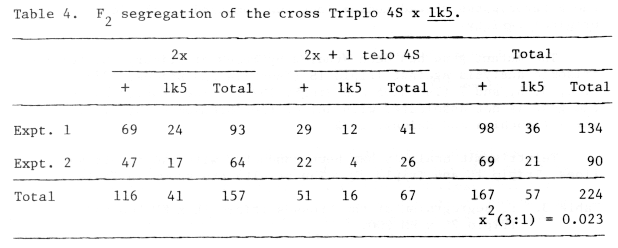
Analysis of lk5: The recessive short awn gene lk5 was first described by Litzenberger and Green (1951) with the cross-over value of 6.9 with Kk alleles on chromosome 4. Later, the symbol lk6 was suggested by Nilan (1964). However, in view of the fact that the gene lk4 of Litzenberger and Green (1951) (lk5 by Nilan, 1964) was proved to be allelic to lk2 (Eslick and Hockett, 1967), the present author suggests that the original symbol lk5 be retained as the official symbol for this gene. The gene has never been located in the linkage map. The gene lk5 (lk6) was tested for its allelic relation with another short awned mutant, KM 218 and Swedish ari-c2 with the result that the short awned mutant, KM 218 and ari-c2 were allelic to lk5 (lk6) (Tsuchiya, 1974). The trisomic analysis showed that the gene lk5 was associated with chromosome 4 (Triplo 4) (Tsuchiya and Haines, 1975). In this study the gene lk5 was studied by means of telotrisomic analysis with Triplo 4S. As shown in Table 4, the results showed that the gene lk5 is not associated with the same arm as the genes K and f9.
Table 4. F2 segregation of the cross Triplo 4S x lk5.
These results are summarized as follows:
References:
Eslick, R.F. and E.A. Hockett. 1967. Allelism for awn length, lk2 in barley (Hordeum species). Crop Sci. 7:266-277.
Hanson, W.D. 1952. An interpretation of the observed amount of recombination in interchange heterozygotes in barley. Genetics 37:90-100.
----- and H.H. Kramer. 1949. The genetic analysis of two chromosome interchanges in barley from F2 data. Genetics 34:687-800.
Litzenburger, S.C. and J.M. Green. 1951. Inheritance of awns in barley. Agron. J. 43:117-123.
Nilan, R.A. 1964. The cytology and genetics of barley. 1951-1962. Monograph Suppl. 3 Res. Studies Vol. 32, No. 1, Washington State Univ. Press.
Singh, R.J. and T. Tsuchiya. 1974. Further information on telosomic analysis in barley. BGN 4:66-69.
Tsuchiya, T. 1962. Radiation breeding in two-rowed barley. Seiken Ziho (Rep. Kihara Inst. Biol. Res.) 14:21-34.
----- 1972. Cytogenetics of telotrisomics in barley. BGN 2:93-98.
----- 1974. Allelic relationships of genes for short awned mutants in barley. BGN 4:80-85.
-----, T. Fachan and T.E. Haus. 1976. Primary trisomic analysis of lg3 (light green) and f3 chlorina in barley. BGN 6:82-84.
-----, and R.L. Haines. 1975. Trisomic analysis of nine mutant genes in barley. BGN 5:67-70.
----- and D.W. Robertson. 1971. Association of a chlorina mutant with chromosome 3. BGN 1:64-65.
----- and R.J. Singh. 1972. Trisomic analysis of a short awned mutant in barley. BGN 2:99.
----- 1973. Further information on telotrisomic analysis in barley. BGN 3:75-78.
{kind=link}
{kind=link}
{kind=link}
{kind=link}