II. 35. Analysis of the first mitotic divisions in germinating seeds of barley.
M. Murata (1), E. E. Roos (2), and T. Tsuchiya (3), Department of Agronomy, Colorado State University, Fort Collins, Colorado 80523 (1, 3) and National Seed Storage Laboratory, USDA, Fort Collins, Colorado 80523(2)
Chromosomal aberrations induced by various kinds of mutagens such as irradiation, chemical agents, or seed aging have mainly been observed at the first mitotic cycle (first metaphase and/or anaphase) in root- or shoot-tips of germinating seeds. The initiation of the first mitosis is influenced by several factors, particularly temperature. Cell division is most active and root and shoot length increase most rapidly at the optimum temperature. Therefore, the beginning of the first mitotic divisions can be estimated from temperature, period of germination, and root or shoot length.
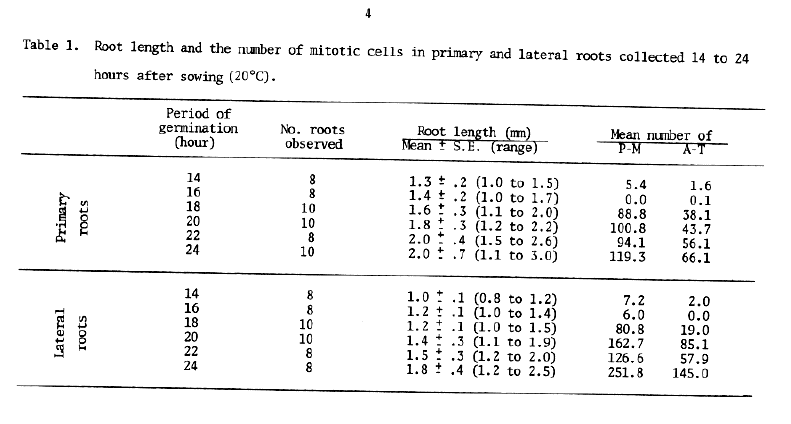
The time of the first mitotic division in root-tip cells was investigated as a prelude to further studies on aged seed. Seeds of Hordeum vulgare cv. Himalaya were germinated on top of filter paper in petri dishes in a constant temperature incubator set at 20°C. Ten seeds were collected at 2-hour intervals from 14 to 24 hours after sowing and fixed in acetic acid-alcohol (3 ethanol: 1 glacial acetic acid). The primary and one of the lateral roots were separately transferred into acetocarmine staining solution (0.8% acetocarmine in 45% acetic acid) after measuring their lengths. Squash preparations were made in 45% acetic acid using about 0.5 mm of the tip of each root. The number of cells per root showing late prophase to late telophase were scored and classified into two groups: 1) late prophase to late metaphase (P-M), and 2) early anaphase to late telophase (A-T) (Table 1).
Root length and the number of mitotic divisions at each collection period are mean values of 8-10 seeds. Few cell divisions were observed at 14 and 16 hours after planting. Both P-M and A-T increased markedly in primary roots collected at 18 hours; thereafter the cell divisions became more active as the germination period increased. A similar pattern was observed in lateral roots. Although the lateral roots were definitely shorter than the primary root in the same seed, no large difference in the number of divided cells was observed between primary and lateral roots. The results obtained suggest that the first mitosis of primary and lateral roots occurs almost simultaneously about 16-18 hours after beginning of germination at 20°C.
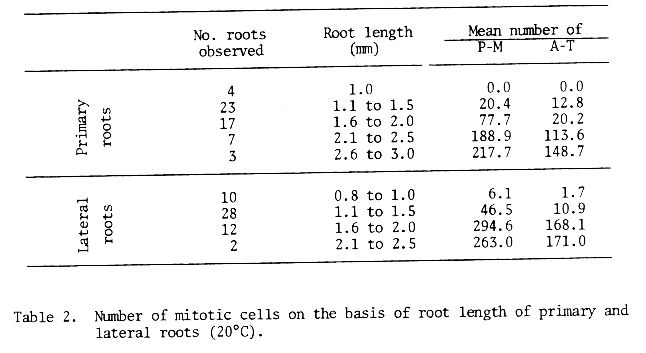
However, root growth varied considerably and ranged from 1.1 to 3.0 mm in primary roots collected at 24 hours. Same shorter roots did not have any dividing cells, whereas other longer roots had more than 400 divisions. Ignoring the period of germination, the average number of dividing cells was calculated on the basis of root length (Table 2.) These data indicate that the dividing cells were found first in primary roots about 1.5 mm in length and in lateral roots about 1.0 mm long. Thus, it is suggested that root length should be used as the indicator of the first mitosis, since the number of dividing cells increased with increased root length.
Table 2. Number of mitotic cells on the basis of root length of primary and lateral roots (20°C).
It has been reported that the first mitotic divisions in barley root-tips begin about 24 hours after germination at 24-26°C (Wolff and Luippold, 1957). In contrast, our data show that the first mitotic divisions began at 16-18 hours after planting at 20°C. Similar results were obtained by Sakurai and Matsumura (1961) who reported that the first mitosis began 16 hours after sowing at room temperature (22-25°C) when the primary and lateral roots had elongated to 1.7 and 1.3 mm, respectively. The differences between their results and our results seem to be caused by the differences of materials and environmental conditions such as temperature and moisture content of the seeds.
According to Künzel (1971) treatment of seeds with chemical mutagens caused a pronounced delay of the first mitotic divisions in both root and shoot meristems. In order to investigate chromosomal aberrations at the first mitotic cycle, any delay in mitosis caused by mutagens or seed aging should be taken into consideration. From these results we can determine the length of any delay in mitosis as a result of seed aging.
References:
Künzel, G. 1971. The ratio of chemically induced chromosome aberrations to gene mutations in barley. Mutation Res. 12:397-409.
Sakurai, N. and S. Matsumura. 1961. On the relation of chromosomal aberrations to gamma-ray dosage an dose rate in barley. Seiken Ziho 12: 61-69. (Japanese with English summary)
Wolff, S. and H. E. Luippold. 1957. Inaccuracy of anaphase bridges as measure of radiation-induced nuclear damage. Nature 179: 208-209.
{kind=link}
{kind=link}