II. 32. Inheritance of the shrunken endosperm character, sex3c, of Bomi Riso mutant 1508 and its association with lysine content.
S. E. Ullrich and R. F. Eslick, Dept. of Plant and Soil Science, Montana State University, Bozeman, Montana 59715.
Shrunken Endosperm Inheritance
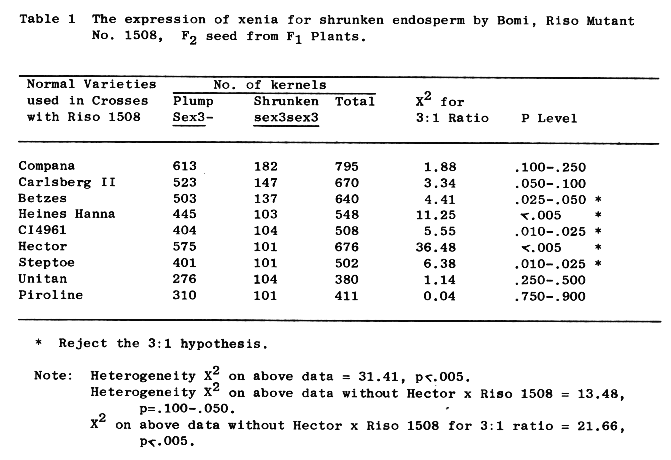
The kernels of Riso 1508 have a characteristic dimple on the dorsal or germ side of the seed in our environments. Presented in Table 1 are data concerning the occurrence of shrunken seed on F1 plants (F2 seed). The data is from crosses of Riso 1508 with nine other varieties. The shrunken endosperm trait is recessive, as indicated by an acceptable fit to a 3:1 ratio for some of the crosses, and this is in agreement with Doll's report (l973) that the high lysine character was inherited as a simple recessive gene.
Since we are reporting on the shrunken endosperm characteristic of Riso 1508, and this trait has shown xenia in all first crosses, we propose that the trait be designated sex3c.
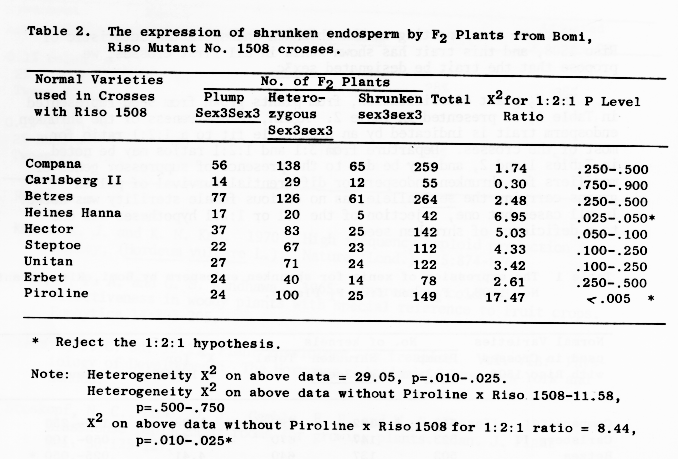
The F2 plant classification, from plants grown from seed represented in Table 1, is presented in Table 2. Again, recessiveness of the shrunken endosperm trait is indicated by an acceptable fit to a 1:2:1 ratio for most of the crosses. Departure from 3:1 and 1:2:1 ratios may be noted in Tables 1 and 2, and may be due to the presence of suppressor genes or modifiers for shrunken endosperm or differential survival of pollen grains carrying the sex3 allele, as no obvious female sterility was noted. In all cases but one, rejection of the 3:1 or 1:2:1 hypotheses was due to a deficiency of shrunken seed.
Table 2. The expression of shrunken endosperm by F2 Plants from Bomi, Riso Mutant No. 1508 crosses.
Shrunken Endosperm sex3 sex3 Linkage
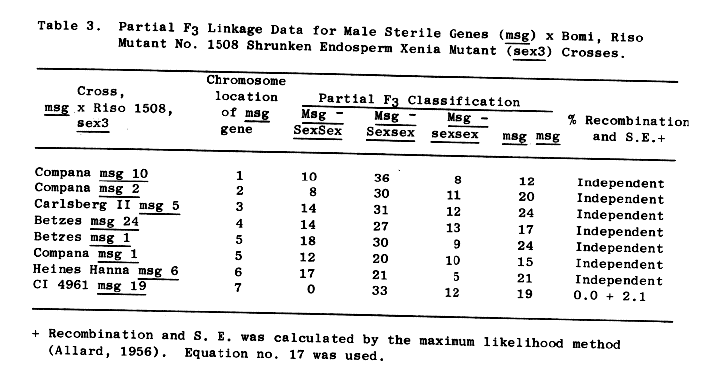
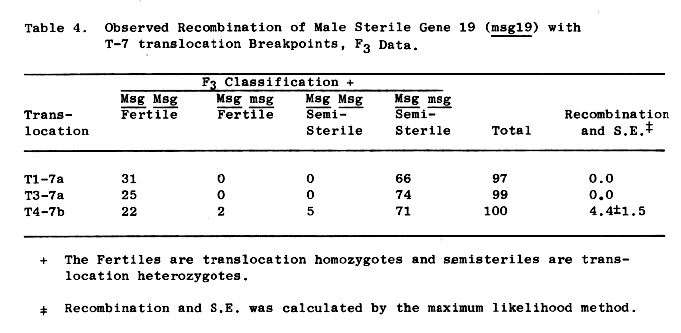
Independence is indicated between sex3 and male sterile genes, msg, on chromosomes 1 thru 6, Table 3. Male sterile 19, msg19, according to Eslick (1971) and the updated data of Table 4, is located on chromosome 7. With the numbers observed, nearly complete linkage between msg19 and sex3 is indicated. The coupled sex3 msg19 genotype was not recovered.
Table 4. Observed Recombination of Male Sterile Gene 19 (msg19) with T-7 translocation Breakpoints, F3 Data.
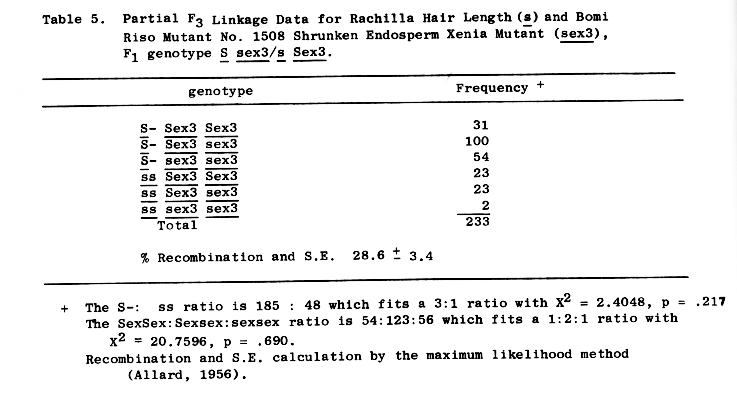
Linkage was also found between the short rachilla hair gene, (s),and sex3. Partial F3 linkage data shows 28.6 + 3.4 percent recombination of the two genes, Table 5.
The short-haired rachilla gene, (s), is usually assigned to the long arm of chromosome 7 (BGN 6:111). K. Karlsson (1972) has indicated the high lysine gene, lys1, of Hiproly is probably 8 units distal to s in the same arm. We have generally considered msg19 to be near the centromere of chromosome 7. The results reported would indicate that the sex3 gene of Riso 1508 is probably near the centromere of chromosome 7. This does not necessarily conflict with the findings of Muench et al. {1976) concerning the independence of Riso 1508's high lysine character and lysl on chromosome 7.
Lysine Association with sex3 sex3
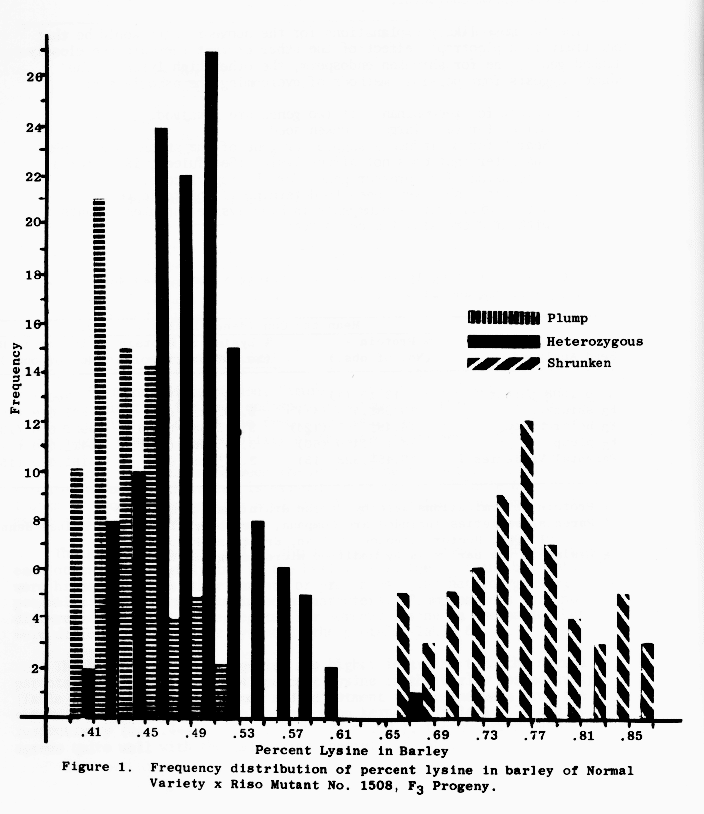
The frequency distribution, Figure 1., suggests a distinct percent lysine distribution for each endosperm genotype, and this is confirmed by a heterogeneity X2 test of the frequency distributions of the plump and heterozygous classes. Probability of the same frequency is <.001.
A close association between the high lysine character and the shrunken endosperm character is evident in Figure 1 and Table 6. Figure 1 data is from F3 progeny (F3 seed from F2 plants) from the same crosses as presented in Table 2.
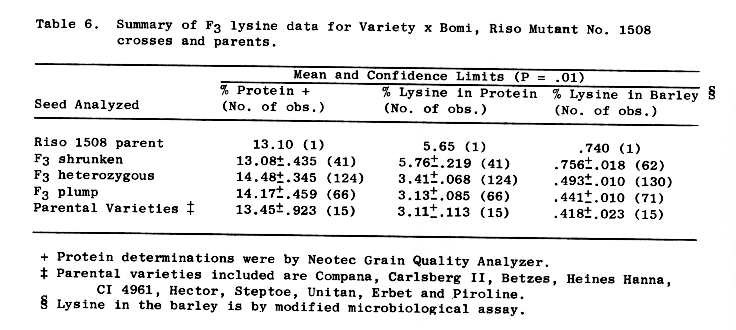
Table 6. Summary of F3 lysine data for Variety x Bomi, Riso Mutant No. 1508 crosses and parents.
The Riso 1508 parent and F3 shrunken progeny were not different from each other in percent lysine in the barley or in the protein, but both were higher than the other parents or progenies. The heterozygous F3 progenies were intermediate in both characters. Plump parents were not different from F3 plump segregates for percent lysine in the barley but were different for percent lysine in the protein, Table 6.
The heterozygous F3 progenies are higher in lysine than the normal parents, .493 percent vs. .441 percent lysine in the barley respectively. (Table 6). If xenia is assumed and adjustment for lysine made from the observed frequencies of plump and shrunken kernels, Table 1, .79 and .21 respectively (eg. .441 x .79 + .756 x .21 = .507), the observed .493 agrees quite well with the calculated theoretical .507.
It follows that the normal lysine characteristic, in agreement with Doll's report (1973), expresses complete dominance. In addition, the high lysine character also expresses xenia in the same manner as the shrunken endosperm character.
The two most likely explanations for the above results would be that one trait is a pleotropic effect of the other or that they are two closely linked genes, one for shrunken endosperm, the other high lysine. The above suggests four possible methods of overcoming the association:
1. Select for recombinants if two genes are involved.
2. Select for very large shrunken seed.
3. Search for or induce a suppressor gene of the shrunken endosperm character that does not affect lysine (See Tuleen, 1968, for a discussion of suppressor genes for chlorophyl color.)
4. Intercross shrunken genes to determine if epistatic gene action will restore plumpness in high lysine shrunken mutants without decreasing lysine content.
References:
Allard, R. W. 1956. Formulas and tables to facilitate the calculation of recombination values in heredity. Hilgardia 24:235-279.
Doll, H. 1973. Inheritance of the high-lysine character of a barley mutant. Hereditas 74:293-294.
Eslick, R. F. 1971. Balanced male steriles and dominant pre-flowering selective genes for use in hybrid seed production. In Barley Genetics II, R. A. Nilan, ed., Wash. State Univ. Press, Pullman. pp. 292-297.
Karlsson, K. E. 1972. Linkage studies on a gene for high lysine content in Hiproly barley. BGN 2:34-36.
Muench, S. R., Lejeune, A. J., Nilan, R. A., and A. Kleinhofs. 1976. Evidence for two independent high lysine genes in barley. Crop Sci. 16:283-285. (and BGN 5:31-32).
Tuleen, N. A. 1968. Genetic investigation of suppressors of a chloroplast mutation in Hordeum vulgare. Genetics 59:44-55.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}