II.4. Male sterile mutants of barley. IV. Different fertility levels of msg9ci (cv. Vantage), an ecoclinal response.
H. Ahokas and E. A. Hockett. Department of Genetics, University of Helsinki, P. Rautatiekatu 13, FIN-00100 Helsinki 10, Finland and ARS, USDA, Plant & Soil Science Department, Montana State University, Bozeman, Montana 59715, U.S.A.
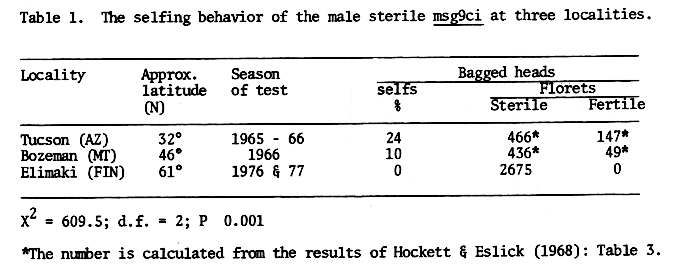
The mutant msg9ci (cv Vantage, CI 7324, Male sterile stock No. 57) was reported to be partialiy self fertile, 10% at Bozeman, and 24% at Tucson (Hockett & Eslick 1968). When tested in Finland in 1976 and 1977, the mutant was found to be completely male sterile. The compiled results are given in Table 1. The differences in the ability to self are highly significant, and thus there is also a significant contingency (C=0.373).
Table 1. The selfing behavior of the male sterile msg9ci at three localities.
The above results could be explained by different photoperiodic conditions. The planting occurred at Tucson in December, at Bozeman and Southern Finland in May. The approximate daylight plus the civil twilight during the early 2.5 months' vegetative period of the season vary at Tucson between 11 - 12.5h, at Bozeman 16 - 17h, and at Elimaki 19.5 - 24h. The light periods were adopted from tables of Anonymous (1945).
A rough assessment of the temperature conditions during the vegetative period at the three localities can be done with the Climatological Data (Arizona vol. 70 & vol. 71; Montana vol. 69; Kuukausikatsaus Suomen ilmastoon vol. 69 & vol. 70, data for Anjalankoski). The temperatures of the early season at the three testing localities don't suggest any correlation to the selfing behavior. Thus, though several important environmental factors have been left out, the present results suggest that the photoperiod is the main ecological factor determining the level of fertility in msg9ci with the genetic background of cv. Vantage. The genetic background will evidently interfere, however (Ahokas 19772.
The suggested photoperiodic response is opposite to the results with the normal fertile barley for spike fertility (Gustafsson & Dormling 1972), and for male fertility (Batch & Morgan 1974).
The tapetum membrane and the deposition of the exine are abnormal in the msg9ci anthers, though this is not necessarily the primary attribute of the male sterilizing gene. The vascular elements of the mutant stamen filament are evidently adequate.
Partial male steriles with a completely sterile response under certain conditions could be useful for hybrid production. A medium or high partial fertility at one locality would make the increase of the seed parent stock economic for hybrid production at another.
References:
Ahokas, H. 1977. Male sterile mutants of barley. V. Tests of cytoplasmic restoration. Cereal Res. Comm. 4:397-400.
Anonymous 1945. Tables of sunrise, sunset, and twilight. Suppl. to the American Ephemeris 1946, U.S. Naval Ohservatory, Washington D.C.
Batch, J. J. & Morgan, D. G. 1974. Male sterility induced in barley by photcperiod. Nature 250:165-167.
Gustafsson, A. & Dormling, I. 1972. Dominance and overdominance in phytotron analysis of monohybrid barley. Hereditas 70:185-216.
Hockett, E. A. & Eslick, R. F. 1968. Genetic male sterility in barley. I. Nonallelic genes. Crop Sci. 8:218-220.
{kind=link}