II.l9 Identification by Giemsa staining of the barley chromosomes and their arms.
Ib Linde-Laursen, Danish Atomic Energy Commission Research Establishment Risø, DK-4000 Roskilde, Denmark.
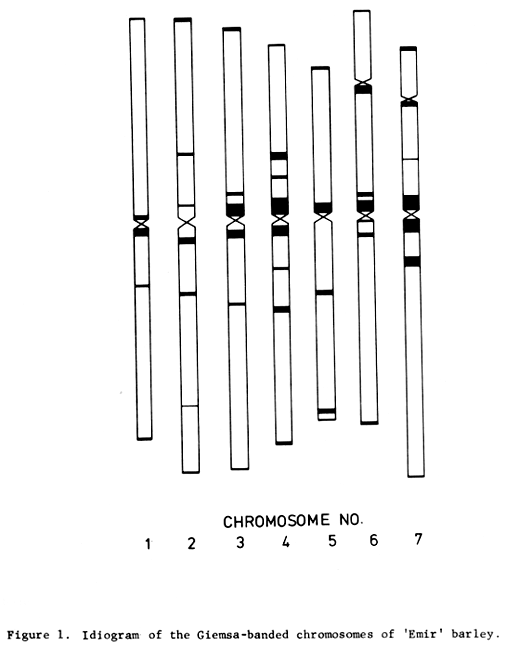
Each of the chromosomes and chromosome arms of the barley variety 'Emir' has its distinctive Giemsa banding pattern. Giemsa banding of the chromosomes of a number of barley varieties has shown their banding patterns to be modifications of a common basic pattern. It is thus possible to identify each barley chromosome with its banding pattern even though plant materials of different origins are used.
Usually only barley chromosomes 5, 6 and 7 are recognizable by conventional cytological techniques. Their Giemsa banding patterns were identified earlier (Linde-Laursen, 1975) (cf. Figure 1). For the identification of the Giemsa banding patterns of barley chromosomes 1, 2, 3, and 4 and of their arms, root tip metaphases of trisomics for chromosomes 1, 2, 3, and 4, and of telotrisomics for chromosome arms 1L, 1S, 2L, 2S, 3S, and 4S (Tsuchiya, 1974) were Giemsa stained. Further, unstained cells of disomic barley were photographed by phase contrast optics, and arm lengths of the somatic metaphase chromosomes, later identified by Giemsa staining, were measured from the photographic negatives.
Figure 1. Idiogram of the Giemsa-banded chromosomes of 'Emir' barley.
The results of the stainings and measurements were:
Trisomics. In each line trisomic for chromosomes 1, 2, 3, and 4, respectively, three chromosomes showed an identical Giemsa banding pattern that, on the whole, agreed with that of the earlier, provisionally designated chromosome nos 1, 2, 3, and 4, respectively, of the barley variety 'Emir' (Linde-Laursen, 1975).
Telotrisomics. 1) Triplo 2L, 2S, 4S. The Giemsa banding patterns of the telocentric chromosomes 2L, 2S, and 4S agreed with those of the longer and the shorter arm of chromosome 2 and with that of the shorter arm of chromosome 4, respectively.
2) Triplo 3S. The Giemsa banding pattern of the telocentric chromosome 3S was invariably found to correspond to the banding pattern of the longer arm of chromosome 3, which on an average made up 56% of the total length of the twenty-two chromosome 3's measured. Based on this fact it is proposed to change the designation of the telotrisomic line Triplo 3S to Triplo 3L.
3) Triplo 1L, 1S. Only in three out of the twenty-three chromosomes measured did the Giemsa banding patterns of the telocentric chromosomes 1L and 1S correspond to those of the long and the short arm of chromosome 1, respectively. In one chromosome the two arms were equally long. In nineteen chromosomes the banding pattern of the telocentric chromosome 1L corresponded to that of the shorter arm and the banding pattern of the telocentric 1S to that of the longer arm. These results suggest that the designation of the telotrisomic lines Triplo 1L and Triplo 1S should be reversed. Further, the banding patterns of the arms of chromosome 1 published earlier (Linde-Laursen, 1975) should be reversed as shown in Figure 1.
References:
Linde-Laursen, I. 1975. Giemsa C-banding of the chromosomes of 'Emir' barley. Hereditas 81 285-289.
Tsuchiya, T. 1974. Coordinator's report: Trisomics. BGN 4: 110-111.
{kind=link}