II.7 Male sterile genes on chromosome 1.
R.F. Eslick, Department of Plant & Soil Science, Montana State University, Bozeman, Montana 59715, U.S.A.
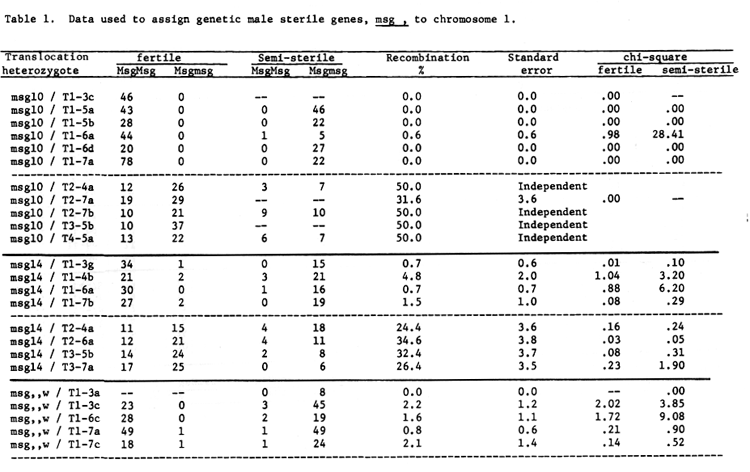
Six non-allelic male sterile genes have been assigned to chromosome 1. Five of the six are considered to be relatively near the centromere and the translocation x male sterile data used to assign the five genes to chromosome 1 are given in table 1. It should be noted that the fertile and semi-steriles were handled as separate populations and maximum likelihood equations used to combine the two populations and calculate recombination values and associated standard errors. The chi-square values given are for goodness of fit of the calculated ratio, based on the reported recombination value, to the observed ratio.
Table 1. Data used to assign genetic male sterile genes, msg , to chromosome 1.
Some of the data of table 1 has previously been presented, Eslick, 1970, namely that for chromosome 1 translocation breakpoints with msg10 and msg14. One may conclude that msg1O is the closest of the 5 genes to the centromere, excepting msg,,k, and that msg22 is probably the furthest from the centromere, and msg14 and msg,,w are probably quite close to the centromere.
Translocations not involving chromosome 1 show a wide range of recombination with the msg genes. Failure to detect independence would seem to be more than coincidence. One would tend to question the validity of recombination values greater than 10 when evaluating data such as those.
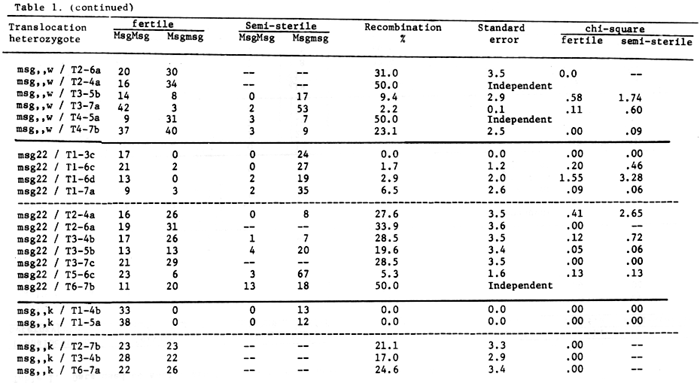
Based on the data of table 2, msg14 may be mapped as below.
Table 2. Data used to position msg14 on chromosome 1.
In another note, Eslick, BGN 6, lk2-n-msg1O-f8 are mapped in that order. The recombination values for msg1O and msg14 and f8 would seem to justify the above gene sequence. By telotrisomic analysis Singh and Tsuchiya, 1974, have placed f8 and f4 in the long arm of chromosome 1. This would be inconsistent with the concept that msg1O does not recombine with the centromere or is very near to the centromere and the gene order would have to be revised to lk2-f4-f8-msg14-msg1O otherwise f8 would be in the opposite arm from f4 and n.
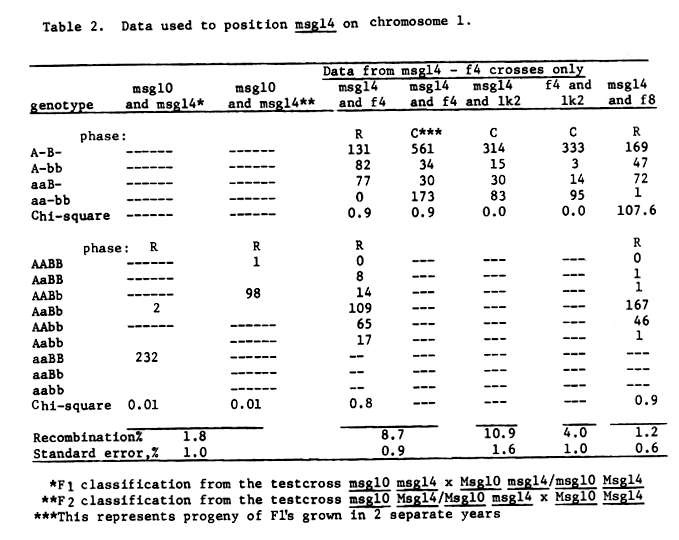
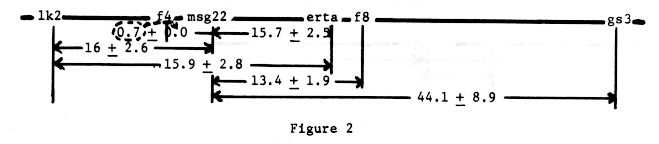
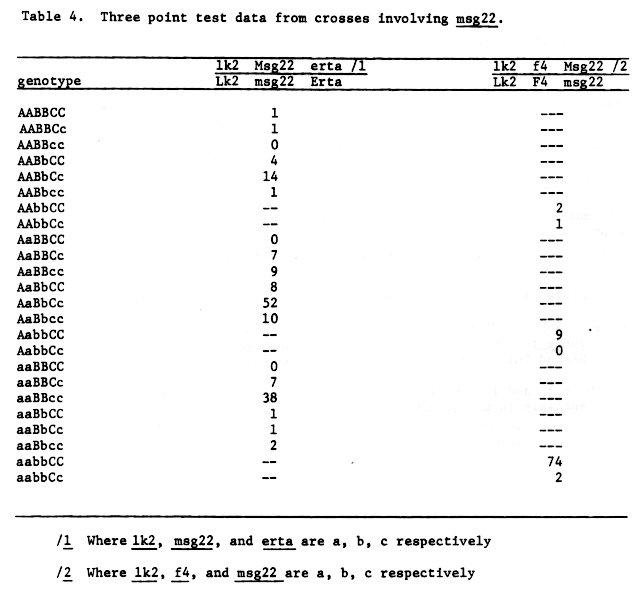
Data from table 3 would justify mapping msg22 very close to f4. The gene orders given in figure 2 are also justified on the basis of three-point data, table 4.
Table 3. Data used to position msg22 on chromosome 1.
Table 4. Three point test data from crosses involving msg22.
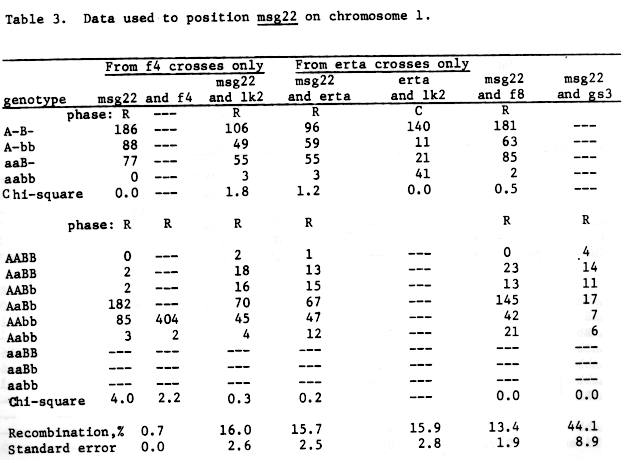
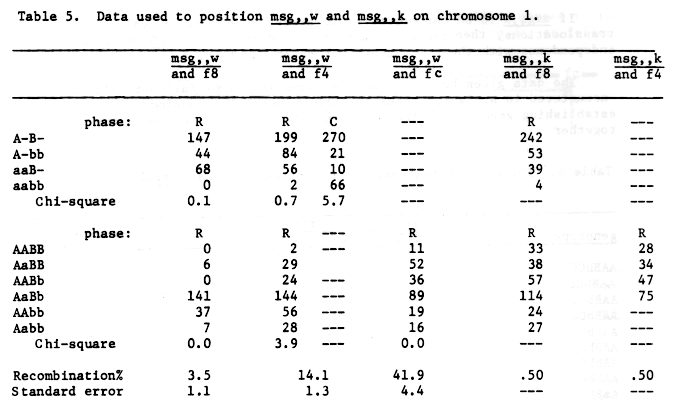
Male sterile msg,,w, table 5, may be mapped as quite close to f8, 3.5% + 1.1. Translocation data would tend to place the gene within l to 2 units of the centromere. It would be distal to msg1O in the long arm if f8 is mapped in the short arm or in the vicinity of msg14 if f8 is mapped in the long arm. Using the available genetic data we would prefer to map as follows:
Table 5. Data used to position msg,,w and msg,,k on chromosome 1.
Note that none of the data for msg,,w is from 3-point crosses.
If msg,,k is on chromosome 1, we prefer recombination values with more translocations, then the gene must be some distance from f4 or f8 as independence with these two genes was observed, table 5.
The data given here demonstrate some of the pitfalls or difficulties encountered in assigning genes to chromosomes with translocations and establishing gene orders from three-point tests when two genes are quite close together.
Literature cited:
Eslick, R.F., 1970 Balanced male sterile and dominant preflowering selective genes for use in hybrid seed production. Barley Genetics II. Proc. Second Int. Barley Genet Symp. pp 292-296.
Singh, R.J. and T. Tsuchiya, 1974 Further information on telotrisomic analysis in barley.BGN 4:66-69.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}