II.34. Further infornation on telotrisomic analysis in barley.
R. S. Singh and T. Tsuchiya, Department of Agronomy, Colorado State University, Fort Collins, Colorado 80521, USA.
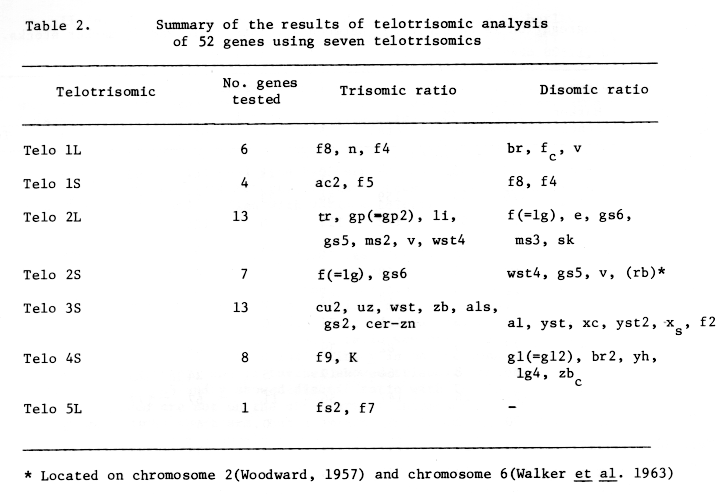
The results of telotrisomic analysis of 34 genes in 7 telotrisomic lines was reported and temporary assignment of these genes on different arms of 5 chromosomes was given (Tsuchiya and Singh, 1973 BGN 3:75-78).
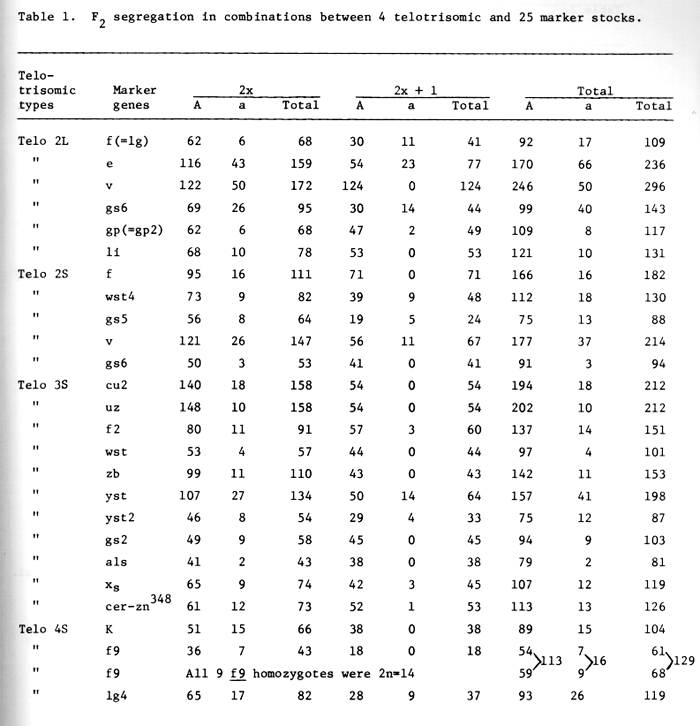
The chromosome number was counted in F2 populations for all cross combinations. No additional gene has been tested with both arms of chromosome 1 after our previous report. Table 1 shows the list of additional genes used in linkage studies with various telotrisomics since the 1973 report.
Since lg and f for chlorina seedlings were found to be allelic (Tsuchiya, 1974 BGN 4:79), the data for both genes were combined together. Further information was obtained for the gene e for elongated glume awn (wide glume) which was reported to be on the long arm of chromosome 2L but showed a disomic ratio with Telo-2L. This indicates that the gene e is on the short arm of chromosome 2 A trisomic ratio was obtained in Vv for two row vs. six rowed spike in Telo-2L indicating v is on the long arm of chromosome 2. The genes gp (=gp2) and li further showed trisomic ratios with Triplo 2L. The genes wst4, gs5 and v showed disomic ratio with Triplo 2S indicating that these genes are not on the short arm of chromosome 2. In the previous map gs5 was on the short arm, but a trisomic ratio was obtained with Triplo 2L. This indicates that gs5 should be moved to long arm from the short arm.
A trisomic ratio was obtained with gs6 in Triplo 2S, indicating that short arm of chromosome 2 carries this gene. This gene has not been located on the linkage map. The frequency of the occurrence of gs6 in the F2 population of Telo 2S x gs6 was very low even in the diploid section, while normal 3:1 ratio was obtained with Triplo 2L (Table 1).
In our previous experiments, genes cu2, wst, and yst2 showed trisomic ratio with telotrisomic for chromosome 3, which was designated as Telo 3L by Tsuchiya and Singh (1973). In this year's experiment the genes cu2, uz, and wst showed trisomic ratio, while f2, yst and yst2 showed disomic ratio.
In our previous note the gene yst2 was reported to show trisomic ratio in telotrisomic analysis with Triplo 3L, because no recessive homozygote was obtained in the trisomic section (Tsuchiya and Singh, 1973). In this experiment, using additional materials, yst2 showed disomic ratio; appreciable number of trisomic recessive homozygotes (4 out of 12) was found, though the overall frequency of recessive homozygotes (13.8%) was much lower than disomic ratio.
This discrepancy may be due to the fact that retested material was kept until maturity and the mutant character appeared late in telotrisomic plants, while the materials were discarded at an early stage in the previous experiment.
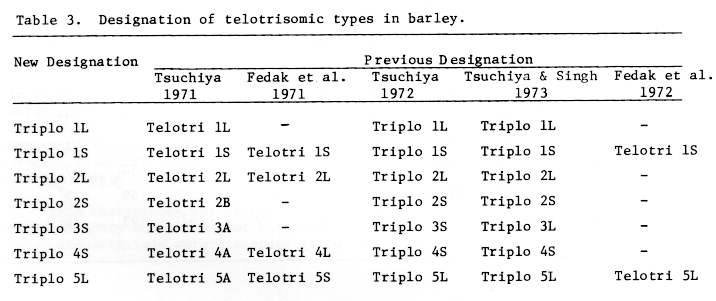
Based on the results shown in Tables 1 and 2 the telotrisomic types are tentatively designated as shown in Table 3 (cf. Tsuchiya, 1974. BGN 4:110).
Table 1. F2 segregation in combinations between 4 telotrisomic and 25 marker stocks.
Table 2. Summary of the results of telotrisomic analysis of 52 genes using seven telotrisomics
Table 3. Designation of telotrisomic types in barley.
Genes for zebra stripe character (zb), als for absent lower laterals, gs2 for glossy sheath, and one Swedish mutant cer-zn348 all showed trisomic ratios in Triplo 3S (formerly 3L). A gene for xantha seedling (xs) which was located at the very end of the long arm of chromosome 3 (Robertson, 1971) shoved disomic ratio confirming that telocentric chromosome is for the short arm of chromosome 3 - (3S) but not for the long arm (3L). The centromere position should be between zb and yst.
Telotrisomic analysis of Kk for hooded vs. awned was completed with Telo 4S and a trisomic ratio was obtained. This indicates that the telocentric chromosome is for the short arm of chromosome 4 (4S).
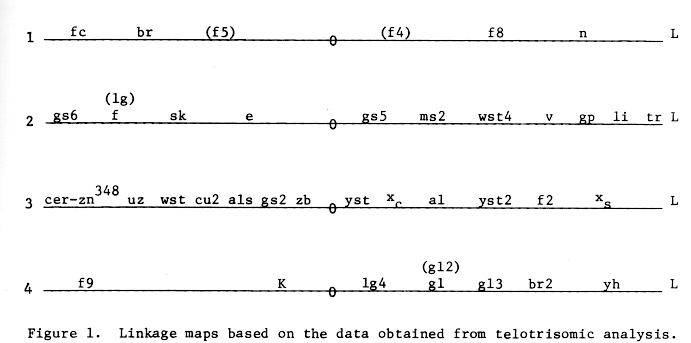
Based on the data obtained from telotrisomic analysis 37(39) genes have been assigned to respective arms of four chromosomes (1 to 4) of barley (Figure 1). The centromere positions are also shown in the map. The order of the genes in the map may not be correct.
References:
Fedak, G., T. Tsuchiya, and S. B. Helgason. 1971. Can. J. Genet. Cytol. 13:760-770.
Fedak, G., T. Tsuchiya and S. B. Helgason. 1972. Can. J. Genet. Cytol. 14:949-957.
Robertson, D. W. 1971. BGN II:220-242.
Tsuchiya, T. 1971. BGN 1:58-60.
Tsuchiya, T. 1972. BGN 2:93-98.
Tsuchiya, T. and R. J. Singh. 1973. BGN 3:75-78.
Tsuchiya, T. 1974. BGN 4:79.
Tsuchiya, T. 1974. BGN 4:110-111.
The research work reported in the papers No. II.33, II.34 (p. 65-69) and II.37 (p. 76) through II.42 (p. 90) by T. Tsuchiya and associates was partly supported by NSF Research Grant GB 30493 and Colorado State University Experiment Station Project (Hatch 8).
{kind=link}
{kind=link}
{kind=link}
{kind=link}