II.7. Positioning of genetic male sterile 5 (msg5) on chromosome 3.
R. F. Eslick, Department of Plant and Soil Science, Montana State University, Bozeman, Montana 59715, USA; and W. L. McProud, formerly graduate assistant, now: North American Plant Breeders, RR2, Brookston, Indiana 47923, USA.
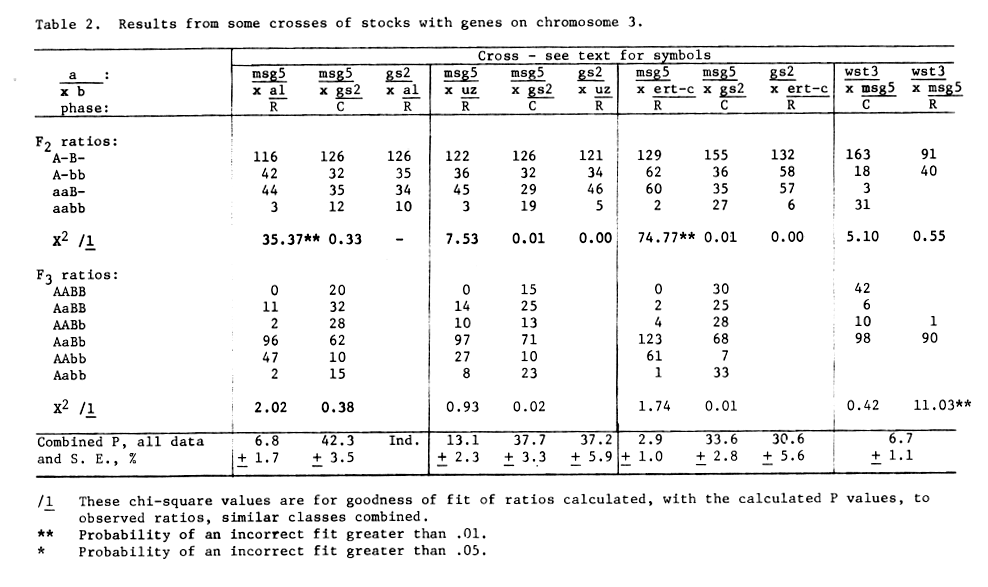
We have accumulated a certain amount of genetic information for mutants assigned to chromosome 3. Although a number of these crosses involved 3 genes a number were so close to msg5 and gs2 was so far removed from msg5 that gene ordering by relative frequency of double cross and single cross gametes was not subject to interpretation.
We have also observed that natural outcrossing in barley may be sufficiently high to account for a certain number of presumably crossover genotypes which may be misleading in determinations when genes are in close proximity.
ALBINO LEMMA, Allb
In 1958, we found a spontaneous albino lemma mutant in the cultivar Liberty. Subsequently we have transferred the gene into Betzes, Compana and Titan, and have used it in a number of crosses.
In allele tests with spring habit derivatives of Takahashi's al al, Russian 82 (winter habit) stock the al al Liberty stock was found to be allelic. The Liberty stock should probably be designated allb allb.
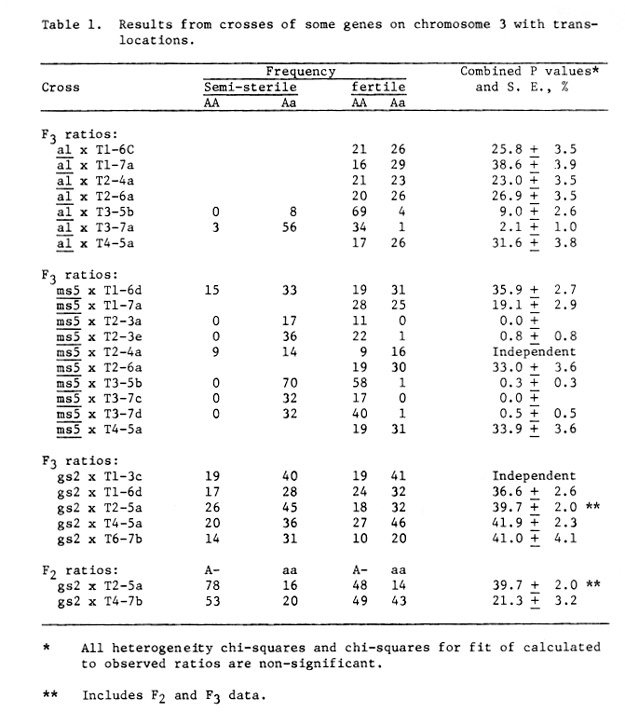
The allb allb gene was assigned to chromosome 3 by the use of translocations. Recombination values of 9.1 + 2.6 percent with the breakpoint of T35b and 2.1 + 1.1 percent recombination with the breakpoint of T37a, Table 1, were obtained. From the translocation data al al is probably in the short arm of chromosome 3, 9.1 + 2.6 percent recombination units from the centromere. The T37a breakpoint is probably in the short arm.
If the genetic data from Table 2 for the gs2 msg5 Al/Gs2 Msg5 al cross is mapped it would appear as follows:
Table 1. Results from crosses of some genes on chromosome 3 with translocations.
Independence of gs2 and al was also reported by Konishi (1973).
UZU DWARF, uz
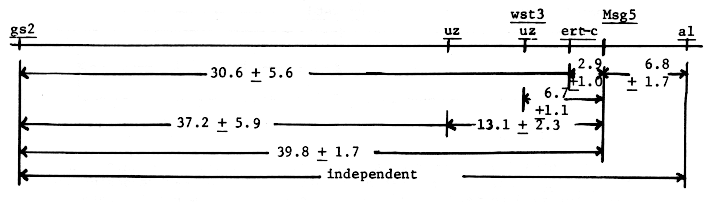
From the cross msg5 gs2 Uz / Msg5 Gs2 uz, Table 2, the three genes may be mapped as follows:
Table 2. Results from some crosses of stocks with
genes on chromosome 3.
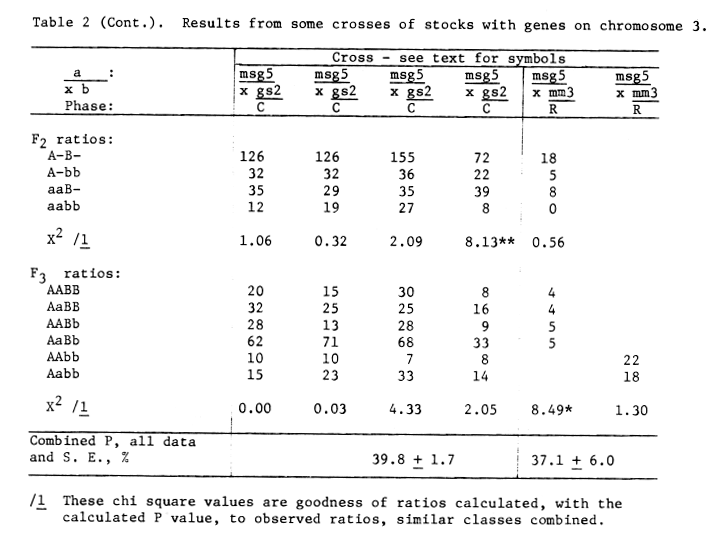
Table 2 (continued). Results from some crosses of
stocks with genes on chromosome 3.
Ramage et al. (1961) reported 14 + 3.1 percent recombination for the T34a breakpoint and uz and 14 + 1.9 percent for uz and the T37b breakpoint. Persson (1969) reported 9.7 + 2.2 percent recombination between uz and the T31j breakpoint. If it is assummed that these translocations are measuring the distance from the centromere to uz, then uz is probably located about 12 units from the centromere in the long arm.
Nonaka (1973) reported 15.8 + 3.3 percent recombination between uz and al. Takahashi and Moriya (1969) reported 8.7 + 1.0 percent recombination and Takahashi and Hayashi (1959) reported 14.2 + 3.2. Perhaps about 11 units from uz to al would be the best estimate.
The observed 37.3 + 5.9 percent recombination for gs2 to uz is in agreement with 37.7 reported by Konishi, (1973).
ERECTOIDES C39, ert-c
Genetic data from the gs2 msg5 Ert-c/Gs2 Msg5 ert-c cross, Table 2, may be mapped as follows:
Persson (1969) reported 22 of 25 translocation crosses to have less than 1 percent recombination with ert-c. T32k had 3.7 + 1.2 percent and T32p had 3.6 + 1.2 percent recombination with ert-c and perhaps estimate the units ert-c is from the centromere. No recombination between ert-c and uz was observed by Persson.
WHITE STREAK 3, wst3
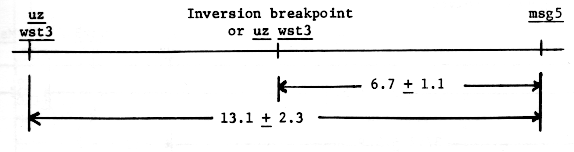
The Uz Wst3 ms5 / uz wst3 Ms5 and uz wst3 ms5 / Uz Wst3 Ms5 crosses were evaluated in F2 and F3. Under our field conditions, classification of wst3 wst3 uz uz plants for fertile and male sterile is difficult and may account for the low number of wst3 wst3 Ms5 _ genotypes observed in the F2 coupling cross, table 2. The other three populations are based on Wst3 - phenotypes. Of the 593 genotypes none represented recombination between wst3 and uz, as also reported by Takahashi and Moriya (1969).
Tsuchiya (1972) reported a paracentric inversion associated with the wst3 uz genes in the Akashinriki stock. The lowered recombination value for msg5 to wst3 uz, 13.1 + 2.3 to 6.7 + 1.1, would indicate the centromere proximal breakpoint of the inversion lies about midway between msg5 and uz. This may also be considered proof of a paracentric rather than pericentric inversion as in a paracentric inversion no msg5 uz wst3 genotypes would have been recovered if a pericentric inversion included wst3 uz.
One hesitates to locate an inversion breakpoint on the basis of these small numbers until an inversion has been shown to include the wst3 uz or uz genes, Tsuchiya and Jensen (1973).
GLOSSY SHEATH 2, gs2
All data available is used to estimate a combined recombination value of gs2 to ms5 of 39.8 + 1.7 percent, Table 2. Glossy Sheath 2 was independent of the breakpoint T31c, Table 1. In a gs2 msg X T2-3g cross the associated male sterile gene was determined to be on chromosome 2, therefore this data is not reported as F3 data cannot be readily interpreted.
MANY NODED DWARF 3, mm3
The mm3 stock used was obtained from G. W. R. Walker and was in the variety Bonus. Walker, et. al. (1963) reported a recombination value of 36.7 + 3.0 percent for mm3 to uz in a coupling cross.
We have had difficulty in distinguishing the male sterile gene in the mm3 genotype. With the inadequate data from table 2 providing the msg5 to mm 3 recombination value of 37.1 + 6.0 percent one could conclude that mm3 is not near the centromere of chromosome 3.
GENETIC MALE STERILE 5, msg5
Genetic male sterile 5 gave little or no recombination with the breakpoints of T32a, T32e, T35b, T37c and T37d, table 1. None of the 3 cross over plants were of the T3 msg5 coupled type and thus could have been produced by natural outcrossing on the F1 plants. These recombination values do not rule out placing msg5 very near the centromere of chromosome 3. The balanced tertiary trisomic msg5 msg5 T37c was obtained and has been added to the world collection.
All of the available data reported here is used below to map the genes involved.
We believe the data supports the placement of msg5 very near the centromere of chromosome 3.
References:
Konishi, T. 1973. Genetic analysis of EMS-induced mutants in barley. BGN 3:28-31.
Nonaka, S. 1973. A new type of cultivar, Mitake, with very few in number, but thick and stiff culms. BGN 3:45-47.
Persson, G. 1969. An attempt to find suitable genetic markers for dense ear loci in barley II. Hereditas 63:1-28.
Ramage, R. T., C. R. Burnham and A. Hagberg. 1961. A summary of translocation studies in barley. Crop Sci 1:277-279.
Takahashi, R. and J. Hayashi. 1959. Linkage studies of the albino lemma character in barley. Ber. Ohara Inst. Landw. Biol. 11:132-140.
Takahashi, R. and I. Mortya. 1969. IV Linkages of four variegated mutants. Ber. Ohara Inst. Landw. Biol. 15:35-46.
Tsuchiya, T. 1972. A case of paracentric inversion affecting genetic behavior of a white streak gene. BGN 2:104-105.
Tsuchiya, T. and D. A. Jensen. 1973. Further results on the allelism relationship between w and wst3. BGN 3:69-70.
Walker, G. W. R., J. Dietrich, R. Miller and K. Kasha. 1963. Recent barley mutants and their linkages II. Genetic data for further mutants. Can. J. Genet. and Cytol. 5:200-219.
{kind=link}
{kind=link}
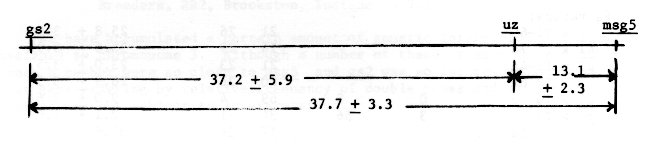{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}