II.26. Confounding the quandary of zygotic DNA*.
L. W. Mericle and R. P. Mericle. Department of Botany and Plant Pathology, Michigan State University, East Lansing, Michigan 48823, U.S.A.
* Auspices of U. S. AEC. Tech. Rpt. C00-1400-43.
Mutagenic treatment of gametes and zygotes has been advocated by a number of barley investigators (e.g., Stadler, 1930; Mericle and Mericle, 1962; Gaul, 1964) with the expectation that all of the generative tissue of the M1 sporophyte would be genetically homogeneous (non-chimeric) for any induced mutation, and hence, provide maximum opportunity for segregation of recessives. This expectation seemed to be realized in the >80% non-chimeric yield obtained in Leonessa barley by Donini et al. (1970) following pollen or egg irradiation. We observed (Mericle and Mericle, 1961) that with sufficiently high radiation level or rate, the entire above-ground portion of an M1 plant of Hannchen barley can arise from only a single cell even as late as the ~500-celled mid-proembryo stage. This suggested that with appropriate manipulation of radiation exposure, the ontogenetic period for achieving non-chimerism can be extended to include most of proembryo development (Mericle and Mericle, 1962).
In contrast to the above, Lundqvist (1964) found that irradiation of mature pollen of Bonus barley almost always produced chimerism for point mutations. Similarly, when we examined more closely the genetic consequences of irradiating zygotes and 2-celled proembryos** of Hannchen barley (Mericle and Mericle, 1967), we were surprised to find, as often as not, a level of genetic chimerism within the generative tissue comparable to that normally exhibited by older proembryos known to possess multicellular meristematic capitals. These results, coupled with cytological implications of cell component partitioning during the first few mitoses after fertilization (Mericle and Mericle, 1969) made it apparent that, in barley, neither pollen, zygotes, nor terminal cells of 2-celled proembryos necessarily behave as expected of typical haploid gametes or diploid meristematic cells according to classical genetic concepts.
[** Considered as single cells since only the terminal cell contributes to the shoot.]
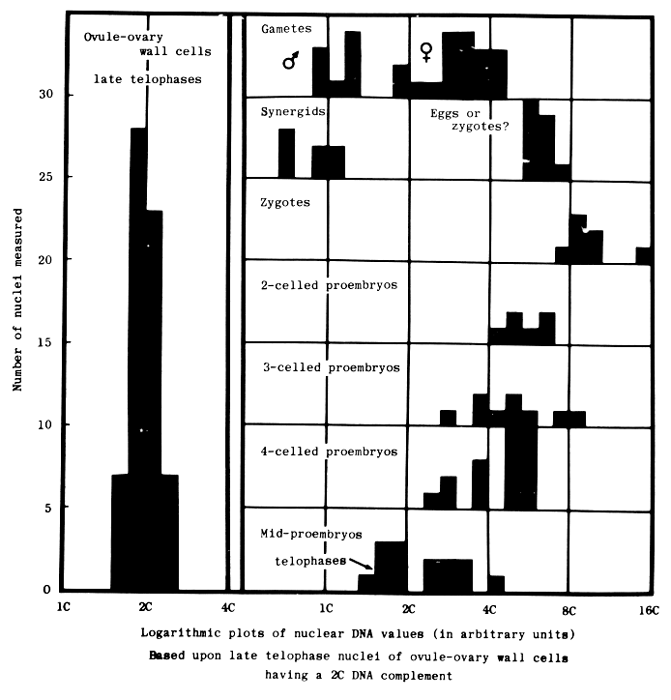
Microspectrophotometry of nuclear DNA content provided a possible explanation. D'Amato et al. (1965) reported 2C complements in barley sperm (cultivar not specified). We found >8C complements in Hannchen zygotes, and persistence of >4C levels through the 4-celled stage -- in the absence of endopolyploidy (Mericle and Mericle, 1970), see Figure 1. D'Amato (personal communication, 1971) suggested our zygote results might be indicative of gene amplification, which they well may be. However, we felt that chromosome polyteny involving all or most of the genome was more likely because of the latent lethality tendencies and chimeric capabilities of the zygotes (Mericle and Mericle, 1970). We concluded (Mericle and Mericle, 1971) that in the absence of strong diplontic selection (causing cell elimination) or severe chromosomal damage, genetic chimerism probably should now be looked upon as the expected consequence of mutation induction in gametes or early proembryos of barley. Instances of nonchimerism can best be accounted for as radiation-induced suppressions of the chimeric potential, although we wonder if cultivar and/or environmental differences might not also be contributing factors.
Studies currently underway in our laboratory were designed to look into the origin of the high DNA content of Hannchen zygotes. We fully expected, on the basis of D'Amato et al.'s 2C value for sperm nuclei, to find DNA contents of at least 2C in Hannchen gametes, and, in view of our >8C zygote values for this cultivar, perhaps as high as 4C. We recently obtained such values for unfertilized eggs, Figure 1. When DNA values appeared not to rise above this range during a 2-3 day post-emasculation period (as one might expect products of gene amplification to do if fertilization were delayed), we become ever more convinced of the existence of polyteny in the zygotes, and gametes as well. But now as we are beginning to examine the DNA content of the male gametes and embryo sac components in the peri-fertilization period, we are completely nonplussed to find only 1C amounts in the male gametes of Hannchen barley, Figure 1. There is no suggestion, as yet, of even 2C values in sperm nuclei which are already within the embryo sac, just prior to fusion. Equally unexpected has been the discovery that, in those instances wherein we are unable to decide on independent, cytological grounds whether an egg has been fertilized or not, the DNA values cluster around 6C, Figure 1.
Whatever the nature of the zygotic DNA may ultimately prove to be, it is already quite clear that its origin is not solely a mathematical addition of DNA complements from the male and female gametes. Considerable DNA synthesis must take place in the immediate post-fertilization period in order for all of the clearly recognizable zygotes to possess at least 8C levels. At present we are becoming more and more inclined to believe that this post-fertilization DNA synthesis involves replication of the male DNA complement, only. Otherwise, the genetic contribution to the sporophyte from the female gamete would be disproportionately much greater. (Do any barley investigators have evidence such might be the case?) If our idea is correct, the 6C cells should represent early post-fertilization stages, immediately subsequent to a first round of male-DNA synthesis, with the 8C zygotes being stages after a second round of synthesis wherein the male and female DNA complements have attained equality. We hope to resolve these questions of post-fertilization synthesis and the nature of the zygotic DNA in the very near future. If evidence continues to accumulate that the zygotic DNA is mostly, or entirely, polytenic replications of the entire genome, then we will very much want to determine whether environment can modify and/or control the pattern of DNA synthesis in the pre-, peri-, and post-fertilization periods, or whether this is a cultivar-specific characteristic. Either could explain the discrepancies in results from gamete irradiation and DNA measurements in male gametes. The possibility of environmental modulation of polyteny could open the door to a better method for deliberate manipulation of chimeric capability in mutation breeding programs than current utilization of high dosages of mutagens, so often accompanied by undesirable levels of sterility.
References:
D'Amato, F., M. Devreux, and G. T. Scarascia-Mugnozza. 1965. The DNA content of the nuclei of the pollen grains in tobacco and barley. Caryologia 18:377-382.
Donini, B., M. Devreux, and G. T. Scarascia-Mugnozza. 1970. Genetic effects of gametophyte irradiations in barley. I. Seedling mutants. Radiat. Bot. 10:79-86.
Gaul, H. 1964. Mutations in plant breeding. Radiat. Bot. 4:155-232.
Lundqvist, U. 1964. Induction of mutations in barley pollen by ultraviolet and X rays. Barley Genetics, Vol. I, Proc. 1st Intern. Bar Genetics Symp., Wageningen. pp. 92-98.
Mericle, L. W. and R. P. Mericle. 1961. Radiosensitivity of developing plant embryos. Brookhaven Symp. Biol. 14:262-286.
Mericle, L. W. and R. P. Mericle. 1962. Mutation induction by proembryo irradiation. Radiat. Bot. 1:195-202.
Mericle, L. W. and R. P. Mericle. 1967. Mutation induction as influenced by developmental stage and age. Erwin Baur Memorial Lectures IV, 1966. Abhandl. Deut. Akad. Wiss. Berlin. pp. 65-77.
Mericle, L. W. and R. P. Mericle. 1969. Cytological consequences of proembryo irradiation. Radiat. Bot. 9:269-282.
Mericle, L. W. and R. P. Mericle. 1970. Nuclear DNA complement in young proembryos of barley. Mut. Res. 10:515-518.
Mericle, R. P. and L. W. Mericle. 1971. Chimerism vs. non-chimerism following irradiation. Genetics 68: s44.
Stadler, L. J. 1930. Some genetic effects of X rays in plants. J. Hered. 21:3-19.
{kind=link}