II.16. Proposal for a new procedure for the preparation of gene maps in barley.
Jens Jensen. Danish Atomic Energy Commission, Research Establishment Risø, DK4000 Roskilde, Denmark.
I should like to suggest the following proposal for the mapping of genes in barley.
The map position 0 should be a gene situated centrally on the chromosome. The 0 position gene should further independently of environmental and genetical background control a character segregating in a perfect Mendelian ratio and be recognizable very early in a generation. The map position of a gene should be given as distance measured in centi-morgans in the direction from the long arm to the short arm of a chromosome (the chromosome is visualized as a number axis, where negative numbers are mainly associated with the long chromosome arm).
The suggestion has the following properties:
1. The 0 position need not be adjusted.
2. The map units = centimorgans are additive. The position of a gene can, therefore, be estimated directly with reference to the 0 position, or its position can be built up by addition of subsequent segments. Further the difference between the positions of two genes on a chromosome map is always the distance in centimorgans between the two genes. For small distances centimorgans and recombination units are practically equal; for long distances a transformation can easily be performed.
3. Placing the 0 position centrally on the chromosome will not give positions with so high values as placing the 0 position at the end of the chromosome, and for that reason the errors of the positions will neither be so large.
4. The conventional way in which to visualize a chromosome diagrammatically is by placing the short arm upwards and the long arm downwards, wherefore the orientation of the gene positions will be in accordance with the normal orientation of a Y-axis in a coordinate system. When placing the gene positions on the X-axis it will be possible to talk about a left and a right chromosome arm as the long and the short arm respectively.
To get an easy and usable transformation of recombination percent to centimorgans I suggest the Kosambi mapping function (see Owen 1950)
cM = centimorgans, ln = natural logarithms, and r = percent recombination. Further, if scM and sr are the standard errors of cM and r respectively, I find
This proposal is made in response to the one given in Barley Genetics Newsletter Vol. 1, (p. 7-8, 1971) which reads "(8) Map positions of gene and chromosome markers should be designated in the following manners: The maps of each arm of each chromosome will have the centromere representing the beginning of each map. The centromere shall represent map position 0. Map position will be described as: the number of recombination units from the centromere, the arm in which the marker is located (S - short and L - long), and the number of the chromosome involved".
This proposal has in my opinion two disadvantages: (1) No method exists at present by which the distance between a gene or another chromosome marker to the centromere can be measured in units which can also be used to measure the distance between genes. (2) As the recombination units, which are required as a measure of map position, are not additive, it will be difficult to construct a gene map, and the position of two genes will tell very little about the distance between them.
My proposal given above does not have these disadvantages.
Reference:
Owen, A. R. G. 1950. The theory of genetical recombination. - In Advances in Genetics 3:117-157.
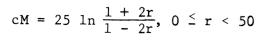{kind=link}
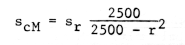{kind=link}