II.14. Cyclic heterozygote advantage at the R/r locus.
S. Jana, B. L. Harvey, and E. T. Thomas. Crop Science Department, University of Saskatchewan, Saskatoon, Sask., Canada.
Substantial heterozygote superiority in fitness was found at the R/r locus for rough versus smooth lemma awn in barley (Jain and Allard, 1960). In a detailed study of Composite Cross V, which was developed from intercrosses among 31 varieties of barley, Jain and Allard observed selective values of x = .82 and y = .68 associated with the RR and rr homozygotes, respectively, where the selective value of the heterozygotes (Rr) was unity. A subsequent study in the Composite Cross XXI, composed of a pool of intercrosses between 6200 strains in the USDA World Collection of barley, revealed little or no selection advantage of the heterozygotes: x = .908 and y = 1.024 (Jain and Marshall, 1967). It is of interest to note that the average outcrossing rate (t) was about 2% in the C.C.V., but was five to seven times higher in C.C. XXI (t = 10 to 15%) due to the inclusion of male sterile strains in the latter population. These results suggest that a balance between natural outcrossing rate and selective value of the heterozygotes may be an important mechanism for maintaining an optimum number of heterozygotes in these genetically variable but predominantly autogamous populations of barley. A verification of this possibility was sought in the work reported herein on the mating system and genotypic fitness at the R/r locus in the bulk propagated sequential generations of a two-parent cross, Husky x Olli.
All generations of the cross were grown at the University of Saskatchewan Farm in Saskatoon using commercial seed rates and standard Saskatchewan agronomic practices. A population of about 15,000 plants was grown in each generation under conditions of severe interplant competition and no conscious human selection. At maturity, the seed was harvested in mass. After thorough mixing, one kilogram of the harvested seed was saved for sowing in the following year and another one-kilogram seed sample was stored at -18°C. A part of the stored seed of each of the eight generations from F4 to F11 was used in the present study. Due to size limitations only the F5, F6, F7, F9, F10 and F11 generations were grown in 1970 and the F8 and F12 in 1971. The plants were grown at a distance of about 8 cm in rows which were 5M long and 30 cms apart. Germination of seed was excellent in each generation (about 98%) and no differential germination was observed.
Husky is shorter, later maturing and higher yielding than Olli. It is a smooth-awned barley (rr), whereas Olli has rough awns (RR). This qualitative difference provided us with an easily recognizable diallelic marker locus, for which population census data could be taken on a large number of plants with accuracy. The census was taken at maturity and plants were classified into rough and smooth awn phenotypic groups. Seeds were collected from each plant separately. In the following year about forty single-plant progenies were grown from the rough-awn phenotypic group for identification of the genotypes RR and Rr. Fifty smooth-awn plant families were also grown in the above manner for an estimation of the outcrossing rate in each generation.
The maximum likelihood estimates of outcrossing rate as outlined by Allard and Workman (1963) were obtained from the proportion of rough-awn plants in the recessive rows. Genotype frequency estimates of sequential generations were used for the maximum likelihood estimates of selective values, x and y, associated with RR and rr genotypes, respectively. Selection was believed to have taken place before the census was taken.
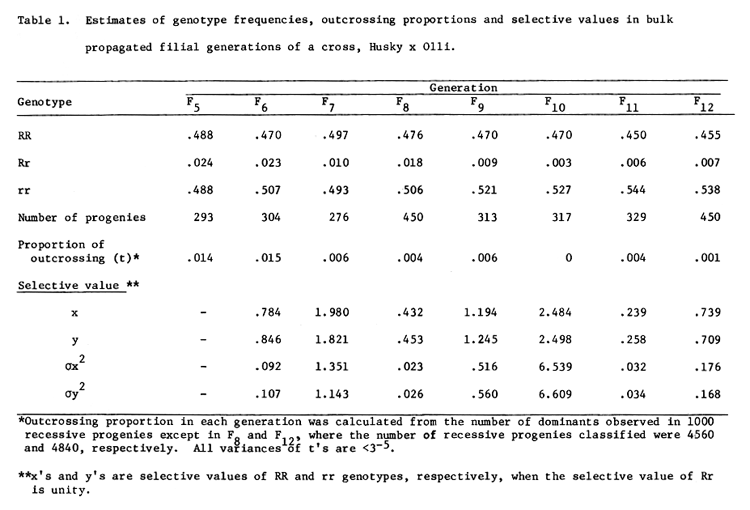
The estimates of relative frequencies of the genotypes, outcrossing rates and selective values are given in Table 1.
The average outcrossing rate at the R/r locus over eight generations was 0.0063 with an empirical standard error of 0.00193. In general, the estimated outcrossing rates were smaller in advanced generations. Because of this appreciable difference, the selective value estimates were based on the outcrossing rate of the preceding generation and not on the average outcrossing rate of 0.0063.
The frequency of RR and rr genotypes were remarkably similar in all eight generations indicating equal fitness of the homozygotes. Thus, selective value estimates of the two homozygotes were very similar in magnitude. However, differential fitness of homozygotes and heterozygotes could be detected in all cases. In four out of the seven generations, heterozygotes appeared to be superior, but were followed by generations of sharply reduced selective values. Although a part of the fluctuations is almost certainly attributable to sampling error, a cyclic change in the relative fitness of the heterozygotes is evident.
In order to determine how much of the variation in heterozygote frequency over generations can be attributed to the outcrossing rate of the previous year we estimated the simple regression coefficient of outcrossing rate in one generation on the proportion of heterozygotes in the succeeding generation (both proportions transformed to angular values). This was non-significant thus disproving a linear functional relationship. However, a high and significant product-moment correlation coefficient was found between the proportion of heterozygotes and outcrossing rate of the same generation: 0.874 on row data and 0.869 after angular transformation. Therefore, if we assume that (i) sampling errors were minimal, (ii) the high correlation between t and R is not spurious, and (iii) there was substantial nongenetic control of outcrossing rate, one would conclude that there was a cyclic adjustment of fitness of the genotypes. This would be an efficient mechanism for maintenance of non-trivial equilibrium in a closed population such as the one we investigated.
Considerably enlarged experiments are in progress at the Crop Science Department, U. of S., for a more detailed study in this area.
References:
Allard, R. W., and P. L. Workman. 1963. Population studies in predominantly
self-pollinated species. IV. Seasonal fluctuations in estimated values
of genetic parameters in lima bean populations. Evolution 17:470-480.
Jain, S. K., and R. W. Allard. 1960. Population studies in predominantly self-pollinated species. I. Evidence for heterozygote advantage in a closed population of barley. Proc. Nat. Acad. Sci. 46:1373-1377.
Jain, S. K., and D. R. Marshall. 1967. Genetic changes in a barley population analysed in terms of some life cycle components of selection. Genetica 38:355-370.
{kind=link}