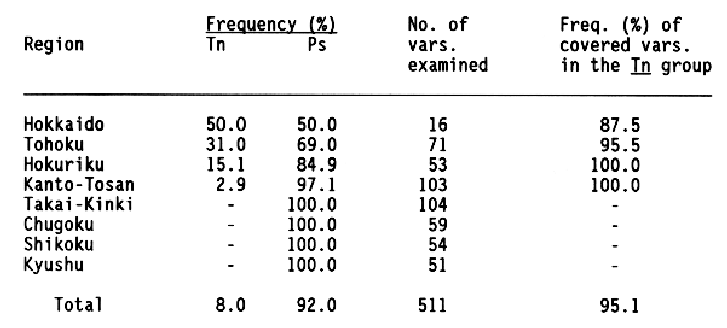
Two alleles, Tn and Ps, were detected at the Pgd2 locus for phosphogluconate dehydrogenase in the World Barley Collection preserved at the Barley Germplasm Center of Okayama University. The Ps allele was frequently found through the world, while the other allele, Tn, was distributed in southwestern Asia, Turkey, and Europe. The Tn allele was also detected in Japanese six-rowed varieties with low frequency (8%). These varieties were grown only in northern parts of Japan, mainly Hokkaido, Tohoku, and Hokuriku regions, and most of the varieties were covered barley, as shown in Table 1.
Among the six-rowed covered varieties grown in the northern regions of Japan, the following differences were observed between two varietal groups carrying the Tn and Ps alleles, respectively. As indicated in Table 2, about 70% of the Tn varieties were resistant to Japanese major races (I, IV, and IX) of powdery mildew, whereas only 15% of the Ps varieties were resistant. And, more than a half of the Ps varieties were susceptible to the races I and IV. The difference is not due to a linkage relationship between the Pgd2 locus and a locus for mildew resistance. These characters are inherited independently even though both of the loci are located on chromosome 5 (Jensen, 1988).
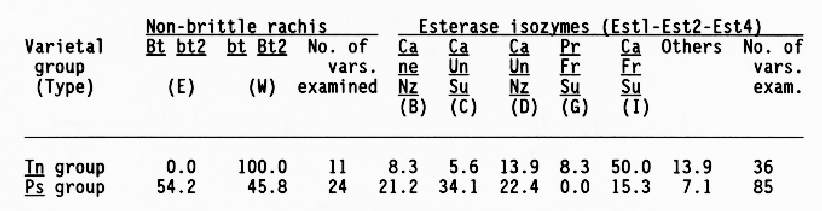
Furthermore, differences were found in genotypes for non-brittle rachis and esterase isozymes. As shown in Table 3, all of the Tn varieties possessed the genotype of bt Bt2 for the non-brittle rachis, which was abundantly detected in the Occidental regions including Turkey and Europe. Meanwhile, a half of the Ps varieties were of the Occidental type (bt Bt2), and the rest were of the Oriental type (Bt bt2) for the non-brittle rachis. The two loci, bt and bt2, are tightly linked, and located on the short arm of chromosome 3 (Takahashi and Hayashi, 1964). In the six-rowed covered varieties of North Japan, five major genotypes (B to D, G and I) for esterase isozymes were found, which consisted of allelic combinations at Est1, Est2, and Est4 loci on the long a chromosome 3 (Konishi and Matsuura, 1987). Out of these genotypes, type I was detected in a half of the Tn varieties. The genotype was frequently found in Turkey and Europe (Konishi, 1988).
From these results, it may be concluded that such differences between the Tn and Ps varietal groups are probably caused by founder effects. Because, it is well-known that some six-rowed covered varieties had been introduced from Europe, and that they were established as Japanese cultivars in the northern regions of Japan. These varieties are characterized by the Tn allele, together with the bt Bt2 genotype for non-brittle rachis and type I of esterase isozymes.
References:
Jensen, J. 1988. Coordinator's report: chromosome 5. BGN 18:61-63.
Konishi, T. 1988. Genetic differentiation and geographical distribution of barley. Crop Genetic Resources of East Asia:237-243.
Konishi, T., and S. Matsuura. 1987. Linkage analysis of Est4 locus for esterase isozyme-4 in barley. BGN 17:68-70.
Takahashi, R., and J. Hayashi. 1964. Linkage study of two complementary genes for brittle rachis in barley. Ber. Ohara Inst. landw. Biol. 12:99-105.