
Barley yellow mosaic virus (BaYMV), which can cause considerable yield losses in malting barley in Southeast Asia and in winter barley in Europe, can only be controlled by resistance breeding. Two sources of resistance genes are used in German and Japanese breeding programs. Some German varieties, such as Franka, Sonate, and Diana, carry a recessive resistant gene originated from the landrace Ragusa (Friedt et al., 1985). The second source of resistance is the Chinese spring barley Mushinchiang 3, called Mokusekko 3 in Japan (Takahashi et al., 1973), which was transferred to spring barley types like Res yml in Japan. The source of resistance originating from Ragusa seems to have been partly overcome (Huth, 1989). Therefore, it is essential to introduce additional germplasm into the breeding material to widen the genetic basis of resistence using the resistance source of Japanese spring barley varieties.
Meanwhile, the doubled haploid (DH) technique is routinely used in practical barley breeding programs. The main advantage of this method is the rapid and complete homozygosity of the offspring, so that phenotype selection for quantitatively and particularly for qualitatively inherited characters is much easier and therefore breeding is more efficient. The central questions, especially for polygenically inherited traits, is: can DHs be used more economically in an F1 generation or in later generations, e.g., a preselected F2 or F3, or should the haploid step be combined with other breeding techniques?
In the case of introduction of BaYMV resistance from distantly related genotypes, such as Japanese cultivars or wild species, into modern German varieties, this was investigated in more detail. The line Res yml was used as starting material for the introduction of new resistant genes into German winter barley varieties. Because Res yml is distantly related to European winter barley material, there are alternative strategies possible for the introduction of this character using haploid techniques. One possibility is a conventional selection program up to the F3 or F4. Here, several rounds of recombination can take place and plants carrying undesirable characters can be eliminated, thus increase the chance of obtaining more valuable DH lines. On the other hand, such a preselection lasts at least 2 years and selection, even for qualitatively inherited characters, is often uncertain. Consequently, with this wide cross, it was thought advantageous to start the haploidization from F1 hybrids, to select immediately in the A1 or, at the latest, in the A2 and to backcross to an optimized new variety, resulting in a new F1.
A prerequisite for the acceptance of this new procedure has been the improved efficiency in the production of DHs per desirable genotype. In barley, under optimal conditions the regeneration frequency of doubled haploid green plants reaches 5-10% based on the number of anthers cultured (Kuhlmann and Foroughi-Wehr, 1989). Table 1 shows the in vitro reaction from F1 crosses following several haploid steps.
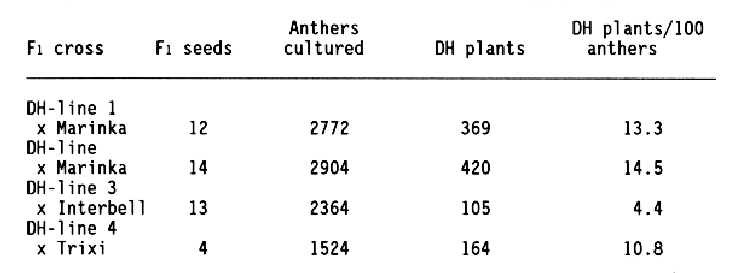
This backcrossing procedure combines recurrent selection with haploid cycles, it is called "recurrent selection alternating with haploid steps" (Foroughi-Wehr and Wenzel, 1990). The model in Fig. 1 dmonstrates this procedure and Table 2 summarizes the results.
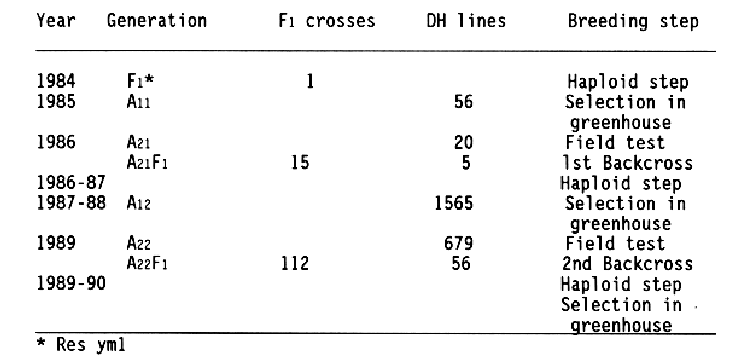
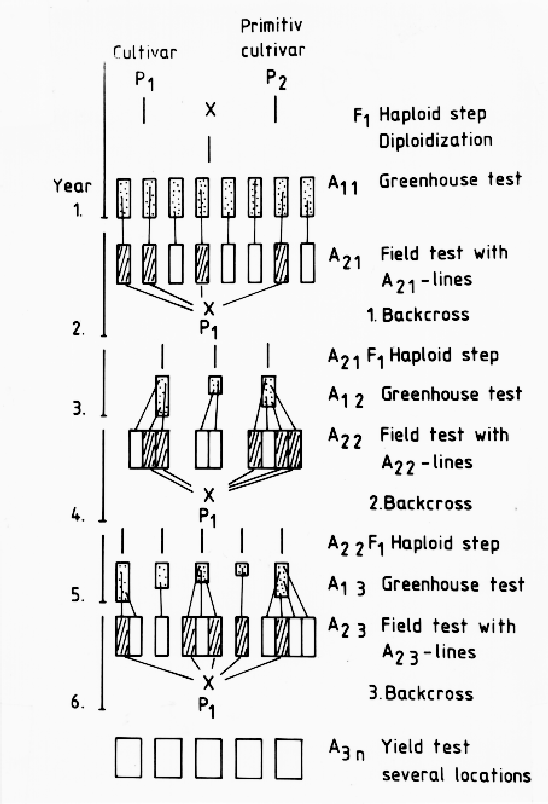
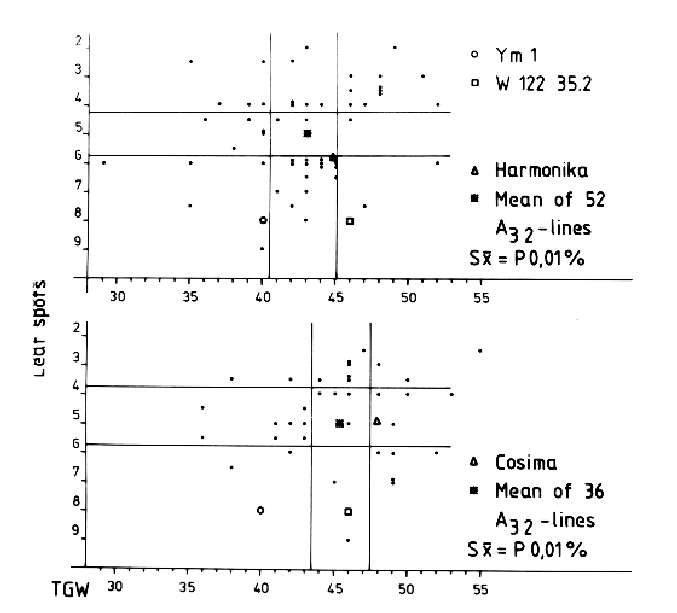
First, only a few DH lines were selected from the F1 (yml x Igri), because most of them carried negative characteristics from the nonadapted Japanese parent, such as lack of chilling resistance, very weak straw, and high susceptibility to leaf diseases. The five best A21 lines were backcrossed with ten leading German vareities. From the F1s about 1500 DHs were regenerated as A12 lines and selected in the greenhouse. After selfing, the A22 lines were additionally screened in the field. The second backcross was carried out with 56 DH lines followed by the third haploid step resulting in the A13 generation. Field tests will give the answer as to whether the developed resistant lines are already close enough to German cultivars or whether an additional cycle is necessary to achieve acceptable lines. Qualitatively inherited characters can be evaluated very soon because of the rapid homozygosity of the offspring. The combination of BaYMV resistance with other important characters between distantly related genotypes within 6-7 years demonstrates the time gains by using the recurrent selection alternating with haploid steps (Table 2). It can be demonstrated that using haploids increases the efficiency of incorporation of mono- and polygenically inherited traits. The leaf diseases and 1000-grain weight (TGW) were recorded and evaluated in DH lines of a first backcross (A22). The starting material was a cross between the high-yielding, BaYMV-susceptible, German winter barley variety Igri and the Japanese spring barley line Res yml. From the first F1, one DH line (W122 35.2) was selected in A21 with chilling resistance and virus resistance. This line was backcrossed to the two leading German varieties Cosima and Harmonika. The BaYMV-resistant DH lines (A22) from the F1 hybrids of the crosses A21 x Cosima and A21 x Harmonika were grown in the field and investigated together with the starting material. The BaYMV resistant line W122 35.2 was higher yielding than the parent Res yml, but resistance against leaf diseases was relatively low. The varieties Cosima and Harmonika were more resistant and their TGW was higher. The DH progeny of the two crosses expressed a tremendous variability for both characters. There were lines with a combination of all the negative characters of the parents, low TGW, and high susceptibility. Some lines still carried the negative characters from Res yml, but there were also DH lines that responded better for both characters than the best parent. From these results it can be concluded that even only one opportunity for genetic recombination is sufficient for a broad spectrum of variability. In the future, recurrent selection alternating with haploid steps could be even more efficient if a positive selection pressure is possible in the A1, e.g., with molecular markers for BaYMV resistance. Such a strategy would speed up the breeding process even more than demonstrated with the present examples, and an early combination of resistance against different diseases will be possible.
References:
Foroughi-Wehr, B., and G. Wenzel. 1990. Recurrent selection alternating with haploid steps - a rapid breeding procedure for combining agronomic traits in inbreeders. Theor. Appl. Genet. 80:564-568.
Friedt, W., W. Huth, H. Mielke, and S. Zuchner. 1985. Resistenztrager gegen Barley Yellow Mosaikvirus. Nachrichtenbl. Deut. Prlanzenschutzd. 37:129-135.
Huth, W. 1989. Ein weiterer Stamm des Barley Yellow Mosaic Virus (BaYMV) gefundes. Nachrichtenbl. Deut. Pflanzenschutzd. 41:6-7.
Kuhlmann, U., and B. Foroughi-Wehr. 1989. Production of doubled haploid lines in frequencies sufficient for barley breeding programs. Plant Cell. Rep. 8:78-81.
Takahashi, R., J. Hayashi, T. Inouye, I. Moriya, and C. Hirao. 1973. Studies on resistance to yellow mosaic disease in barley. I. Tests for varietal reactions and genetic analysis of resistance to the disease. Ber. Ohara Inst. 16:1-17.