11. 28. The micro gametic lethal gene "let", its potential use in a hybrid barley system, and the discovery of the haploidy initiator gene "hap" ("let", "hap" with hybrid barley).
L. Lehmann, Cereal Department, Svalof AB, S-268 00 Svalov, Sweden, G. Hagberg, Swedish University of Agricultural Sciences, Department of Plant Breeding, 5268 00 Svalov, Sweden and P. Hagberg, Biology Department, Svalof AB, S-268 00 Svalov, Sweden "R"
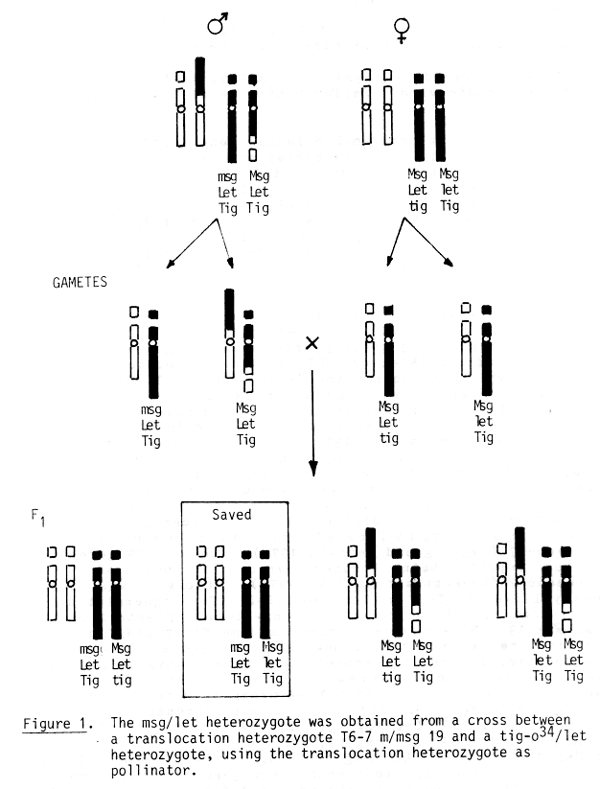
In 1975, let/tig o34 genetic line was received from Diter von Wettstein. This line segregated approximately 1:1 for tigrina versus normally green plants. The two genes are closely linked in repulsion (Nielsen, O.F. 1974). The objective was to find out on which chromosome the "let" gene was located and possibly if it was linked to a known male sterile gene in repulsion. This would facilitate its use in male sterile production for hybrid barley according to the "haplo-viable" system diagramed by Ramage (1976). The first step was crossing the let/tig o34 onto a translocation tester set. Nearly all the pollen should carry the tig o34 gene except for possible escapes or recombinants. The tig o34 gene was used in the linkage studies because it should to easy to read and a gametic lethal would not be involved to disturb the normal segregation. The tig o34 gene was found to be located near the centromere of chromosome 7 (Hagberg and Lehmann 1978). The msg 19 gene had also been reported being located near the centromere of chromosome 7 (Hockett and Eslick, 1970). This gene was also closely linked to T6-7 m (Karlsson, B-A 1976, Hagberg et al. 1978). Root tips from progeny of semi-sterile plants from the cross msg 19 x T6-7 m were examined as the first step in obtaining the let Msg 19/Let msg 19 heterozygote. The 6-7 m translocation is easy to recognize. The heterozygous plants were maintained and at the same time the line let/tig o34 was sown. The heterozygous plants T6-7 m/msg 19 were crossed onto let/tig o34 (Figure 1). This cross should produce four genotypes: N tig o34 Let Msg 19/N Tig o34 Let msg 19, N Tig o34 let Msg 19/N Tig o34 Let msg 19, N Tig o34 let Msg 19/T Tig 034 Let Msg 19, N Tig o34 Let Msg 19/T Tig o34 Let Msg 19. In 1978, the seeds from the described cross were germinated and root tips examined. There were 54 plants examined from five crosses. Of these, 32 had the normal karyotype with 4 plants being tigrina.
These tigrina plants indicated that there was some self-pollination. There were 14 (NT) translocation heterozygotes. These plants were discarded. The most surprising part was that we found 8 haploid plants, of which three were tigrina. None of the haploids had the translocation. Some of the haploids besides the tigrina plants were weak and died. This was the first time we had found so many haploids at one time. On rare occasions we have found haploid plants in the progeny of trisomics. The karyotype of one of the trisomic progenies has been published by Hagberg (1977). The isolation from the parental material let/tig 034 and further research with this haploid initiator gene "hap" have been described by Hagberg and Hagberg (1980, 1981).
Some of the progenies from the normal karyotype were examined cytologically. The first three lines below should have had the genotype N let/N msg 19.
Plant "1". VH78/P16-5a. Seventy seeds were sown of which 66 grew and could be identified. There were 65 diploids and 1 haploid. 65 diploid plants grew to maturity and segregated; 41 fertile plants to 24 sterile plants.
Plant "2". VH78/P16-5b. 70 seeds were sown of which 67 grew and could be identified. All 67 plants were diploid. 64 plants grew to maturity and segregated: 36 fertile plants to 28 sterile plants.
Plant "3". VH78/P16-5c. 236 seeds were sown of which 232 grew and could be identified. There were 230 diploids and 2 haploids. 216 diploid plants grew to maturity and segregated: 112 fertile plants to 104 sterile plants.
Plant "4". VH78/K1a. This plant and the next plant should have had the genotype N tig o34/N msg 19. 70 seeds were sown and 67 grew and could be identified. 66 were diploids and 1 haploid. Segregation of mature diploids was 8 tigrina: 38 fertile plants to 18 sterile plants.
Plant "5". VH78/K1b. 70 seeds were sown and 66 grew and could be identified. All 66 were diploids. Segregation of plants was 18 tigrina: 38 fertile plants to 8 sterile plants. If there were close linkage and no other factors were involved, these last two progeny tests should have segregated about: 1 tigrina: 2 fertile: 1 sterile.
In 1979, 29 progenies of Plant "4" were sown. 24 seeds of each progeny was sown. They all segregated for tigrina, fertile, and sterile plants. Of the plants that grew, there were 341 fertile, 161 sterile plants, and 9 plants that were tentatively identified as haploids by their morphological characters. Unfortunately the number of tigrina plants was not noted. The segregation by fertile to sterile plants fit close to the 2:1 ratio.
Seeds from 33 progenies of Plant "1" were sown. There were sown 24 seeds from each progeny. All lines segregated for fertile and sterile plants. The segregation was 376 fertile plants, 349 steriles, and 15 plants that were tentatively identified as haploids by their morphological characters. In order to test the transmission or recombinants fertile plants were crossed onto the male steriles. There were sown nearly 738 seeds from 56 crosses. One cross gave a 1:1 segregation, while another gave only fertiles. The rest of the crosses gave mostly sterile plants. From 54 crosses, the segregation was 17 fertile: 569 sterile plants. This was a higher frequency of fertile plants, than from other pollen lethals that we have previously reported (Lehmann et al. 1979, Lehmann 1981). The percentage of fertiles was considered to be too high to continue the work with the "let" gene and msg 19 as a system to be used in the hybrid barley program.
References:
Hagberg, A. 1977. Vaxtforadling - gron evolution. Pg 47. - Natur och Kultur, Stockholm pp. 153.
Hagberg, A., L. Lehmann and P. Hagberg. 1978. Segmental interchanges in barley. II. Translocations involving chromosome 6 and 7. - A. Pflanzenzuehtg. 8:89-110.
Hagberg, A, and G. Hagberg. 1980. High frequency of spontaneous haploids in the progeny of an induced mutation in barley. -Brief report. - Hereditas 93:341-313.
Hagberg, G, and A. Hagberg. 1981. Haploid initiator gene in barley. - Barley Genetics IV:686-689.
Hagberg, P. and L. Lehmann. 1978. Translocation linkage studies of a tigrina mutant tig-o34. - Barley Genet. Newsl. 8:46-48.
Hockett, E.A. and R.F. Eslick. 1970. Genetic male sterile genes useful in hybrid barley production. - Barley Genetics II:298-307.
Karlsson, B.A. 1976. Studie over kopplingsforhallanden mellan hansterilitetsgener oeh translokationsbrottpunkter i korn. -Inst. Genet., Univ. Lund.
Lehmann, L., P. Hagberg and G. Hagberg. 1979. Pollen lethal induced with sodium azide. - Barley Genet. Newsl. 9:57-58.
Lehmann, L. 1981. Where is hybrid barley today? - Barley Genetics IV:727-777.
Nielsen, O.F. 1974. Macromoleoular physiology of plastids. XII. Tigrina mutants in barley: genetic spectroscopic and structural characterization. - Hereditas 76:269-304.
Ramage, R.T. 1976. Hybrid Barley. - Barley Genetics III:761-770.
{kind=link}