II. 16. Hordeum vulgare autotriploids.
Juliana Balkanjieva, Insitute of Genetics, 1113 Sofia, Bulgaria.
Hybridization between spring barley forms differing in their ploidy level with the aim of producing autotriploids has been studied by Tsuchiya (1958, 1960) paying particular attention to the influence exerted by direction of crossing on seed set and seed viability. Seeds produced as a result of this hybridization were usually smaller and wrinkled, because the course of their tetraploid or pentaploid endosperm development was considerably disturbed. Seed viability was reduced to a great extent and in most cases the seeds germinated with difficulty or not at all, because, as a result of postgamic incompatibility, the embryo either degenerated at an early age or remained undifferentiated. The incompatibility was partially overcome by embryo culture, eliminating the maternal endosperm and cultivating the embryo on artificial nutrient medium.
The present communication presents data about multi-row barley autotriploids produced by embryo culture.
Genotypes characterized by certain morphological traits-type and length of the awn and the spike, used as markers, were selected as parental forms.
Only autotetraploids were chosen for female parents, because the percentage of viable seeds produced by 2x x 4x crosses is very low (Tsuchiya 1958, Balkanjieva, 1983). The autotetraploids Ager and 1241 were developed by colchicine treatment at the Institute of Genetics in Sofia, while Missouri, Riniken spät and Tetra smooth were obtained from the Institute fur Resistenzgenetic, Grunbach - BRD.
The crosses were made in the field. Immediately after pollination the florets were sprayed with a solution of 50 ppm giberellic acid (GA3) and 50 ppm 6- Benzylaminopurin (BAP) in order to stimulate zygote development and to reduce the degree of incompatibility between embryo and endosperm. The twelve-day-old embryos excised from the endosperm were cultivated under controlled conditions on White's medium (1963) with supplements after Atanassov (unpublished data) as follows: vitamins 1 ml/1, Fe chelate - 5 ml/1, sucrose - 15 g/1, glucose - 15 g/1, yeast extract - 200 mg/1, casein hydrolysate - 200 mg/1, giberellic acid - 0.01 mg/1, kinetin - 0.05 mg/1, ANA - 0.05 mg/1.
The chromosome number of the plants was determined after Feulgen squash preparations.
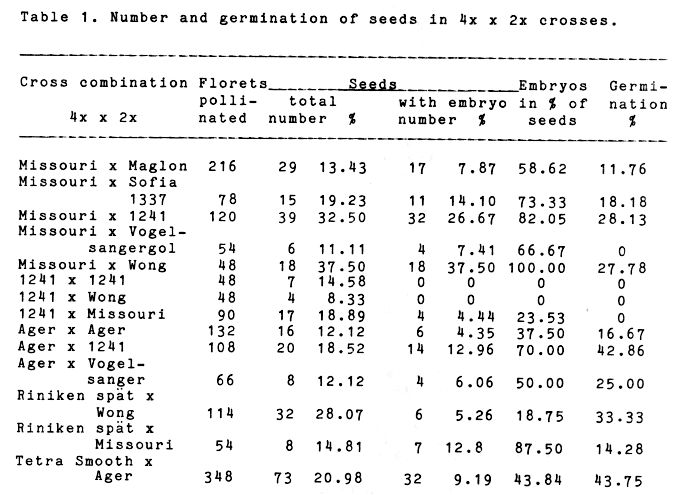
All crosses set seeds, but after eliminating the endosperm it became evident that many of the seeds had no embryo. With the exception of the combination Missouri (4x) x Wong (2x) immature seeds with embryos were always less than total number of seed set, while 1241 (4x) x 1241 (2x) and 1241 (4x) x Wong (2x) produced no seed set (Table 1). This fact indicates that the values for hybrid and non-hybrid seeds produced reflect only partially and incompletely the degree of incompatibility between tetraploid and diploid forms. Besides that, they do not reveal the presence of embryos, combining the diploid and haploid gametes from which a triploid plant is going to develop. Therefore, the presence of an embryo reflects more accurately the degree of its incompatibility with the endosperm and depending on viability, it either germinates or dies. Immature or mature seeds without an embryo are non-viable and no plant can grow out of them. Lack of embryos in the seed set can be accepted as the main reason for the reported greatly reduced germination of triploid barley seeds (Tsuchiya 1958, 1960), rye seeds (Hakansson and Ellerstrom 1950, Pilch 1978) and seeds of other plant species. It represents the final result of the disturbances due to post gamic incompatibility in the development of triploid embryos and pentaploid or tetraploid endosperm in 4x x 2x and 2x x 4x crosses respectively.
Table 1. Number and germination of seeds in 4x x 2x crosses.
It is of great interest to note that a great number of seed sets produced do not always ensure the same number of embryos. The cross Missouri (4x) x Wong (2x) is characterized by high seed set and embryo values, 37.50% of pollinated florets, but in the cross Riniken spät (4x) x Wong (2x) excised embryos are much less than the immature seeds produced - 5.26% and 28.07% respectively.
Data presented in Table 1 substantiate the assumption that double fertilization and seed set production take a specific course and are of varying extent in each cross. Viable and nonviable seeds are produced, depending on the degree of incompatibility between the triploid embryo (in which two genomes of the female parent and one of the pollinating parent take part) and the pentaploid endosperm (combining four genomes of the female parent's genotype and one of the pollinating parent). While the values of immature seeds show that fertilization has taken its course to a certain extent, the values of immature embryos express the degree of surmounted incompatibility and reveal the practical possibility for producing triploid plants by use of "in vitro" methods. It is of greater significance and more importance for the solution of practical problems, in case contemporary "in vitro" methods are applied, to produce from a small number of pollinated florets not more seed set, but more viable immature embryos. Only 8 immature seeds were produced by the cross Riniken spät (4x) x Missouri (2x), but 87.50% of them were with embryos, while the combination Riniken spät (4x) x Wong (2x) yielded a high number of seed set, but only 18.75% of the seeds had embryos.
Results concerning the number of seed sets produced, their state and potential for developing into plants differ considerably in the individual crosses. Combinations in which one and the same tetraploid female parent takes part are no exception. This is an evidence of the genotypes effect on seed set and seed viability, as well as on barley crosses with closer or more distant Triticinae relatives.
Differences were observed also between the various crosses in size and degree of embryo differentiation, endosperm quantity, structure and colour, seed coat colour and thickness. Seed set produced by crossing the autotetraploid from Missouri with five diploids had hybrid embryos varying in size and differentiation and endosperm of different quantity and structure. The cross Tetra Smooth (4x) x Ager (2x) was distinguished by very large embryos, but many of them were yellowish, soft, with symptoms of an accelerated course of degeneration processes taking place at an early stage. The fruit set of the crosses 1241 (4x) x 1241 (2x) and 1241 (4x) x Wong (2x) were distinguished by a thick seed coat, and a small quantity of or no endosperm. The observed and pointed out difference between hybrid and non-hybrid immature seeds and embryos, obtained from the separate crosses are an indication of the specific influence and interaction between the genotypes at the time of their formation.
A total of 163 embryos were placed to germinate on a nutrient medium. The germination of the individual combinations varied from 0 to 43.75% (Table 1). Observations showed that after 5-6 days the larger embryos develop first their coleoptile and next their rootlets. Some embryos, however, germinated after 3-4 weeks. It can be presumed for the cases with abnormal development, where only leaves or only rootlets were developed, that the 12-day-old embryos had not been fully differentiated or that the degeneration processes had occurred at an earlier age. However, the unfavourable effect of some environmental factors at the time the embryos were excised from the endosperm or at the time of their cultivation on nutrient medium should not be excluded. The triploid plants grown in a glasshouse developed slowly, a number of them perished at different developmental stages. Thirty-seven eutriploids with chromosome number 3x=21 were produced from the 44 germinating embryos.
Comparison of these results with data obtained in the investigations of Tsuchiya (1958) on the hybridization of autotetraploids with diploids in view of producing barley triploids is a difficult task, because the method of embryo culture was not applied. However, the high degree of incompatibility found in the crosses observed in these investigations is confirmed by the present study.
Results obtained indicate that the method of embryo culture can be successfully applied for producing barley autotriploids. Isolation of the embryo from the endosperm and its cultivation on an artificial nutrient medium eliminates the effect of the existing incompatibility between them. The viability of the immature and mature seeds produced depends on the presence of a viable triploid embryos in them.
References:
Balkanjieva, J. 1983. Hybridization between autotetraploid and diploid forms of barley. I. Influence of reciprocal crossing on hybrid seed and viability. Genetics and plant breeding 16:43-51.
Hakansson, A. and S. Ellerstrom. 1950. Seed development after reciprocal crosses between diploid and tetraploid rye. Hereditas 36:256-298.
Pilch, J. 1978. Originating of rye triploids (Secale cereale L.) as initial material for obtaining primary trisomics. Part I. Genetica Polonica 19:121-136.
Tsuchiya, T. 1958 Studies on cross compatibility of diploid, triploid and tetraploid barleys. I. Seed development in reciprocal crosses between diploids and induced autotetraploids. Bot. Mag., Tokyo 71:48-53.
Tsuchiya, T. 1960. Studies on cross compatibility of diploid, triploid and tetraploid barleys. II. Results of crosses between triploids, diploids and induced autotetraploids. Jap. Jour. Genet. 35:337-343.
White, P.R. 1963. The cultivation of animal and plant cells. 2nd ed.
The Ronald Press, New York.
{kind=link}