II.1. Localization of induced genes for powdery mildew resistance.
G. Robbelen and M.Heun1, Institute of Agronomy and Plant Breeding, Georg-August-University, 3400 Gottingen, Germany, F.R.
1 Present address: Institute of Agronomy and Plant Breeding, TU-Munchen, 8050 Freising-Weihenstephan, Germany F.R.
A wide range of different mutants has been selected by treating seeds of the barley cultivar 'Bomi' with EMS or NaN3 and subsequent selection for changed mildew reaction (Abdel-Hafez and Robbelen 1979; 1980; Kwon et al. 1981; Varghese and Robbelen 1984). Except E292 (see below), all mutations are inherited in a monogenic recessive manner. Some were proven to be allelic with the ml-o gene (Reinhold 1980a,b; Kwon et al. 1980); but most of the induced resistance genes are independently inherited. For evaluating mutagenic techniques in broadening the genetic variation, it was to be analyzed how many loci can be involved in such changes in mildew resistance after mutagenic treatment. This report summarizes our work to localize and pool the induced genes into allelic groups.
Table 1: Segregation ratios in F2 of crosses between marker lines and mutant E61
The following marker genes of chromosomes 4 and 5 (both carrying known resistance genes) have been used for crosses: ml-o, br-2, K, f-9, lk-5 (chr. 4), ned-1a, B and wst-5 (chr. 5). Unfortunately, the spontaneous resistance genes also used in our crosses were not sufficiently expressed to identify clearcut F2 classes. In F1 plants, seed set was shown not to be impaired by meiotic disturbances. Germination of F2 seeds was as high as in the parental lines. This excludes selection after fertilization, i.e. at the zygote level.
B1101 is a partially resistant mutant (Abdel-Hafez and Robbelen 1980). In crosses with known ml-o mutants (M66, SR1, B1012, B1865, N105), the mutated gene of B1101 could be identified to be an ml-o allele. This shows dominant inheritance in crosses with the above mentioned strongly resistant ml-o mutants, but a recessive one with the wild type (Heun and Robbelen 1984). The different phenotypic expression of the new ml-o allele against the earlier ones in identical 'Bomi' background (such as 1012, B1865 and N105) indicates its heteroallelic nature. The typical ml-o necroses are much less apparent, and yields of the mutant are equivalent to the original 'Bomi'.
N182, another partially resistant mutant, is affected by early browning of leaves in the field and consequently lowered yields. The mutated gene of N182 exhibited linkage to four marker genes of chromosome 4 (br-2, K, lk-5 and f-9). According to the estimated recombination values this gene is located in or nearby the M1-g locus (Heun 1984).
E202 shows high resistance except for specific pathotypes; leaves develop large necroses even without infection. F2 segregation data are influenced by gametic selection reducing mutant frequencies. With the appropriate adaptions (Heun 1982) free segregation was found in all crosses with the above mentioned marker genes on chromosomes 4 and 5. Minimal recombination values were calculated which would have been detected in these experiments (Heun 1984). Such distances by which linkage of the mutated gene to the respective marker is excluded, overlap along the total length of chromosorues 4 and 5L. These chromosomes, therefore, do not carry the E202 gene.
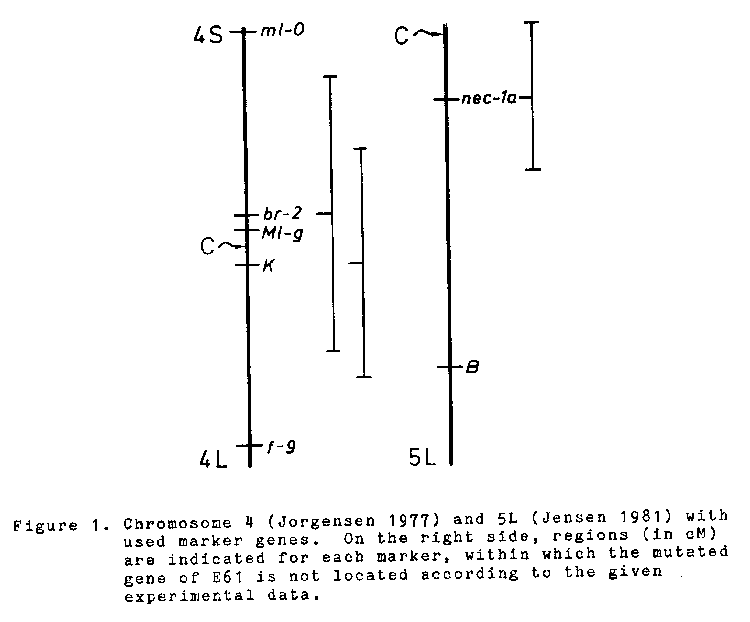
E61 also showing pathotype specificity is not located in or nearby the ml-o locus (Kwon et al. 1981; Reinhold 1980 a,b). F2 ratios of crosses with marker genes are shown in Table 1. Calculating regions where the E61 gene is not located (Figure 1) only the distal end of chromosome 4L remains undetermined.
The conclusion, the E61 as well as the E202 gene is independent from the ml-o and the N192 locus. The same is true for the dominant, partially resistant mutant E292 (with exceptionally poor growth habit) as was shown by crosses with marker genes br-2 and ml-o (Heun 1982). The ml-o and N182 mutants identify two different loci; but the rest of the induced resistance genes (like E202, E61 and E292) investigated genetically so far can only be described as to be inherited independently from the first two loci. For the localization of other of our induced resistance genes the short arm of chromosome 5 carrying most of the spontaneous resistance genes deserves priority, since N182 was located in or nearby the spontaneous M1-g resistance.
References:
Abdel-Hafez, A.-A.G.I. and G. Robbelen. 1979. Differences in partial
resistance of barley to powdery mildew (Erysiphe graminis DC.f.sp. hordei
Marchal) after chemomutagenesis. I. Screening of mutants under field conditions.
Z. Pflanzenzuchtg. 83:321-339.
Abdel-Hafez, A.-A.G.I. and G. Robbelen. 1980. Differences in partial resistance of barley to powdery mildew (Erysiphe graminis DC.f.sp. hordei Marchal) after chemomutagenesis. II. Reaction of mutants to pathotypes. Euphytica 29:755-768.
Heun, M. and G. Robbelen. 1984. Heteroallelism in the ml-o locus of barley. Z. Pflanzenzuchtg. 92:281-288.
Heun, M. 1982. Lokalisierung induzierter Resistenzgene gegen Mehltau bei Gerste. Ph.D. Thesis, Georg-August-Universitat, Gottingen.
Heun, M. 1984. Localization of induced genes of barley for resistance against powdery mildew. Z. Pflanzenzuchtg. 93:158-168.
Jensen, J. 1981. Coordinator's Report: Barley Genetics Newsletter Chromosome 5. 11:87-88.
Jorgensen, J.H. 1977. Location of the ml-o locus on barley chromosome 4. In: Induced Mutations Against Plant Diseases, Proe. Intern. Symp. IAEA/FAO/SIDA, Vienna. 533-549.
Kwon, H.-J., G. Robbelen and M. Heun, 1981. Inheritance and pathotype specificity of mildew resistance of induced barley mutants. In: Barley Genetics IV, University Press, Edinburgh. 441-448.
Reinhold, M. 1980a. Genetische Charakterisierung von Mutanten mit abweichender Mehltaureaktion aus einer Gerstensorte mit mittlerer Resistenz. 1. Quantitative Erbanalyse im Freiland. Z. Pflanzenzuchtg. 84:63-77.
Reinhold, M. 1980b. Genetische Charakterisierung von Mutanten mit abweichender Mehltaureaktion aus einer Gerstensorte mit mittlerer Resistenz. 2. Erbanalyse nach Infektion mit Pathotypen under kontrollierten Bedingugen. Z. Pflanzenzuchtg. 84:89-99.
Varghese, Y.A., and G. Robbelen. 1984. Sectorial chimeras as a proof of the de novo origin of mutants showing partial resistance to powdery mildew in barley. Z. Pflanzenzuchtg. 92:265-280.
{kind=link}
{kind=link}