II. 16. Preliminary evidence of the extent of genetic diversity in parallel populations of Composite Cross Five.
David Luckett* and Abid Sharif*, Department of Genetics, University of Cambridge, Downing Street, Cambridge, CB2 3EH, U.K. "R"
*Present Addresses: CSIRO Division of Plant Industry, Cotton Research Unit, P.O. Box 59, Narrabri, NSW 2390, Australia and Department of Genetics, University of Newcastle, NE1 7RU, UK, respectively.
Recently Luckett (1983), Edwards and Luckett (1983) and Luckett, Sharif and Edwards (1983) have reported studies to determine the possible action of natural selection within 3 parallel populations of Composite Cross Five (CCV) derived from the Californian material and grown in Cambridge, UK. The Cambridge populations are designated I, II and III (Edwards and Luckett, 1983). Polymorphisms at 4 seedling esterase loci and a number of metrical characters have been examined. Here we report preliminary work to establish the level of genetic diversity within the Cambridge CCV populations.
The composite populations can be regarded as heterogeneous mixtures of homozygous and genetically isolated lines, each represented by one or more individuals. If diversity is high then a large number of distinct lines will be present; if low then only a few common lines will predominate.
Inbred lines were isolated at random from the extreme generations (designated '0' and '6' by Edwards and Luckett, 1983) of two of the CCV populations by single-seed descent and selfed in the glasshouse for a further two generations. Replicate individuals of each line from the two population I generations were scored for 5 morphological characters (each controlled by single loci, 4 of which were biallelic), 6 enzyme systems (starch gel electrophoresis of the products of 14 loci, 6 of which were polymorphic) and slab acrylamide gel electrophoresis of the highly variable B and C hordein storage proteins (coded by two multigene complexes) to attempt to unambiguously distinguish as many of the lines as possible. Population III lines were characterized for the same 14 enzyme loci (the same 6 were polymorphic), but not the hordeins, and for only two of the morphological characters (both of which were monomorphic).
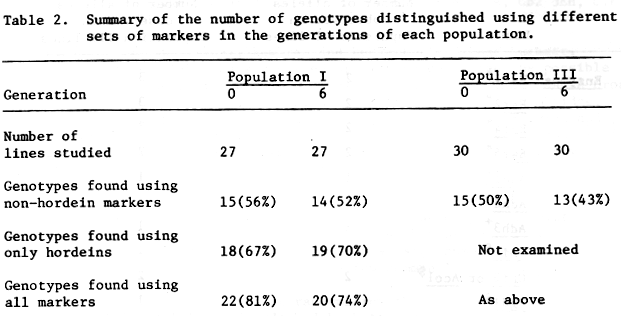
A list of the loci and the alleles detected in lines from each population are given in Table 1. Table 2 shows the number of genotypes determined among the lines of each generation.
Table 1. A list of the loci examined and the number of alleles detected at each.
The hordeins were highly polymorphic (Doll and Brown, 1979; Shewry et al., 1980) and the 33 different overall banding patterns detected were sufficient to distinguish 37 of the 54 lines from population I. Four patterns were found in both generations; the most common genotype was represented by 5 lines and only in generation 6. The additional resolution provided by the other nine polymorphic loci raised the proportion distinguishable to 42/54 (Table 2). Using only non-hordein markers the number of genotypes defined was 29/54. The lines from population III were separated into 28/60 genotypes using only the 6 enzyme polymorphisms (Table 2).
Although the sample size is small the results suggest that the level
of genetic diversity in both populations is high with a large number of
different genotypes present and that the level is not strikingly different
between the generations within populations (Table 2). Doll and Brown (1979),
Muona (1980) and Chai (1977) have examined the frequencies of inbred line
genotypes in other composite barley populations and have all reported high
genetic diversity.
The somewhat surprising conclusion is that selection does not push these heterogeneous populations towards uniformity, even after many generations of strong inbreeding; rather it appears that negative frequency dependent selection may act to provide an advantage in being rare. If this is the case then it may be of value to include such populations in a barley breeding programme; a decision that needs to be considered most carefully.
References:
Brown, A.H.D. 1983. Barley. In 'Isozymes in plant genetics and breeding,
Part B' pp 57-77 eds. S.D. Tanksley and T.J. Orton, Elsevier, Amsterdam.
Chai, M. 1977. The relationship of several characteristics in randomly selected lines of barley composite cross XXI. Dissertation Abstracts International 38:991B.
Doll, H. and A.H.D. Brown. 1979. Hordein variation in wild (Hordeum spontaneum) and cultivated (H. vulgare) barley. Canad. Jour. Genet. Cytol. 21:391-404.
Edwards, K.J.R. and D.J. Luckett. 1983. Hitch-hiking esterase genes in barley. Submitted to Genetics.
Harberd, N.P. and K.J.R. Edwards. 1983. Further studies on the alcohol dehydrogenases in barley: evidence for a third alcohol dehydrogenase locus and data on the effect of an Adhl-null mutation in homozygous and in heterozygous condition. Genetical Research 41:109-116.
Hvid, S. and G. Nielsen. 1977. Esterase isoenzyme variants in barley. Hereditas 87:155-162.
Jain, S.K. and R.W. Allard. 1960. Population studies in predominately self-pollinated species I. Evidence for heterozygote advantage in a closed population of barley. Proc. Natl. Acad. Sci. USA 46:1371-1377.
Kahler, A.L. and R.W. Allard. 1970. Genetics of isozyme variants in barley. I. Esterases. Crop Science 10:444-449.
Luckett, D.J. 1983. Natural selection in genetically heterogeneous barley populations. PhD, University of Cambridge, Cambridge, UK.
Luckett, D.J., A.L. Sharif and K.J.R. Edwards. 1983. Changes in metrical characters in two parallel barley populations. Submitted to Genetics.
Muona, 0. 1982. A Multilocus study of an experimental barley populations. Hereditas 96:247-254.
Shewry, P.R., A.J. Faulks, R.A. Pickering, I.T. Jones, R.A. Finch and B.T. Miflin. 1980. The genetic analysis of barley storage proteins. Heredity 44:383-389.
{kind=link}
{kind=link}