II. 8. Crossability relationships between certain species in the
Hordeae.
R.A. Pickering, Welsh Plant Breeding Station, Plas Gogerddan, Aberystwyth,
Dyfed, SY23 3EB, Wales, U.K.
There have been several reports suggesting a relationship between incompatibility systems in the Hordeae involving wheat, barley, rye and Hordeum bulbosum L. (Fedak and Jui, 1982; Pickering, 1983a; Snape et al., 1979) and other wild species (Thomas et al., 1981). There are similarities in their genetic control and, in some cases, in the manifestation of the incompatibility (pollen tube bursting within stylar tissue).
The dominant gene governing incompatibility of H. vulgare L. cv. Vada with H. bulbosum is known to be linked to the gene conditioning DDT response located on barley chromosome 7 (Pickering, 1983a). This particular chromosome shows some homoeology with wheat chromosomes of group 5 (Islam and Shepherd, 1981) on which two dominant incompatibility genes controlling the incompatibility response with H. bulbosum and rye are located (Snape et al., 1979; Riley and Chapman, 1967).
Since locating the gene in Vada, we have carried out similar investigations using three other barley cultivars possessing incompatibility with H. bulbosum. Recombination values with the DDT gene on chromosome 7 of 12.1% (Mandolin - VDH 479-72), 15.4% (Firlbeck III), and 17.9% (Melody) were obtained compared with 11.2% for Vada. As yet the origin of the incompatibility in these three cultivars has not been identified, as all the parents in their pedigree have proved to be compatible with H. bulbosum, including those related to Vada (e.g. Minerva in Mandolin and Delta in Melody). However, we know that variation for incompatibility with H. bulbosum exists between stocks of the same cultivar (e.g. cv. Universe - Pickering and Morgan, 1978), and the possibility remains that stocks of Minerva and Delta used for breeding Mandolin and Melody were incompatible with H. bulbosum. Firlbeck III is not likely to be related to Vada.
Further evidence in support of a relationship between the incompatibilities of barley x H. bulbosum and those involving wheat x H. bulbosum and wheat and rye was obtained after pollinating four cultivars of barley (Vada, Emir, Golden Promise and Sabarlis) with Chinese Spring wheat. Only on Vada (incompatible with H. bulbosum) was there no seed set, while on the other three cultivars (compatible with H. bulbosum) a range of seed sets was obtained (Table 1).
Table 1. Seed sets on four barley cultivars pollinated with Chinese Spring wheat during May 1983.
Similar crosses have been made by Elbern (1981) using Chinese Spring as both male and female. All hybridizations from which seeds were obtained involved the use of parent cultivars which we have found to be compatible with H. bulbosum. However, no seeds were recorded when cv. Carina was used as either male or female. One of the parents of Carina is Union and this we know possesses a dominant incompatibility gene with H. bulbosum inherited from Firlbeck III. It is likely that Carina is also incompatible with H. bulbosum, and we have since been able to confirm this. Elbern found, after undertaking genetical tests, that one or two dominant genes are present in Carina which control incompatibility with wheat. It will be interesting to determine whether the same dominant genes) in Carina and Vada cause incompatibility with both Chines Spring and H. bulbosum.
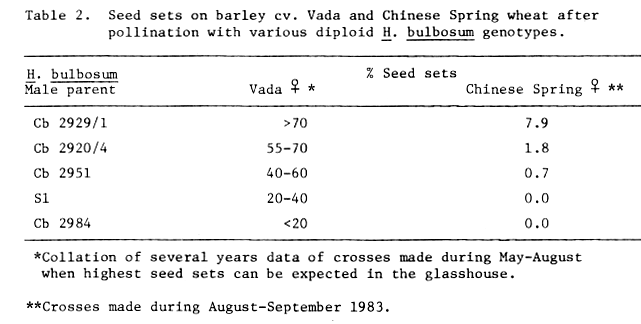
We have also pollinated Chinese Spring with five diploid H. bulbosum genotypes, which give significantly different levels of seed set on Vada (P = 0.05 or less), and the results are presented in Table 2. Although Chinese Spring wheat possesses recessive alleles for incompatibility with H. bulbosum and rye, seed set after pollinating with diploid H. bulbosum is relatively low compared with tetrapolid H. bulbosum (Barclay, 1975) and rye (e.g. Falk and Kasha, 1981). We obtained the highest seed sets on both Vada and Chinese Spring when Cb 2929/1 diploid H. bulbosum was used as pollen donor, and the ranking orders of seed sets show close agreement (Table 2).
The same five H. bulbosum genotypes have been used in 2-component pollen mixtures on three barley cultivars compatible with H. bulbosum. Pollen of one of the components was weighed and mixed with an equal quantity of the other component. In every case, one of the genotypes used in the mixture was Cb 2951 which produces over 80% globular type embryos in crosses with H. vulgare (Pickering, 1983b) and therefore acted as a marker. All the other H. bulbosum genotypes tested produce very low proportions of this type of embryo. Embryos were examined 2-3 weeks after pollination and classified into four categories on their morphology (Pickering, 1983b). The components of the mixture which had effected fertilization could therefore be determined by comparing and analysing statistically the numbers of each type of embryo obtained from the mixture and the components used individually. In pollen mixtures of Cb 2951 with a) Cb 2929/1 and b) Cb 2920/4, fertilization was accomplished by Cb 2929/1 and Cb 2920/4 respectively, whereas in mixtures of Cb 2951 with c) S1 and d) Cb 2984, Cb 2951 achieved preferential fertilization. Therefore, the ranking orders of seed sets on Vada and Chinese Spring using these five H. bulbosum genotypes correspond with their performance on barley cultivars compatible with H. bulbosum. There were no significant differences in pollen grain numbers per unit weight of pollen between Cb 2951, Cb 2929/1 and Cb 2920/4, but there were significantly greater numbers of grains of S1 and Cb 2984 compared with Cb 2951. Differences in total numbers of pollen grains in each component of the mixtures could not therefore account for the preferential fertilization. In order to detect differences between H. bulbosum genotypes in pollen tube growth rates, pollinations of three compatible H. vulgare cultivars were carried out using each H. bulbosum genotype individually. Ovaries were fixed at 10-120 minutes after pollination, stained and examined using fluorescence microscopy. It was not possible to observe any differences in pollen tube lengths between pollinators, but more critical assessments might be helpful. Pollen grain viability and germination were not estimated, but control seed sets obtained on the same three H. vulgare cultivars using each component of the mixtures separately never fell below 90%.
From the data presented, we can conclude that certain H. bulbosum selections will outperform others in compatible and incompatible hybridizations, and that several incompatibility systems in the Hordeae are closely related. It must be remembered however, that seed setting varies considerably according to technique, environment and genotype and is not an ideal indicator of crossability. Nevertheless, it is clear that certain cross combinations, especially those involving rye, are more successful.
References:
Barclay, I.R. 1975. High frequencies of haploid production in wheat
(Triticum aestivum) by chromosome elimination. Nature 256:410-411.
Elbern, B. 1981. Moglichkeiten der Bastardierung zwischen Gerste, Weizen and Roggen. PhD thesis; Friedrich-Wilhelms Univ., Bonn.
Falk, D.E. and K.J. Kasha. 1981. Comparison of the crossability of rye (Secale cereale) and Hordeum bulbosum onto wheat (Triticum aestivum). Can. J. Genet. Cytol. 23:81-88.
Fedak, G. and P.Y. Jui. 1982. Chromosome of Chinese Spring wheat carrying genes for crossability with Betzes barley. Can. J. Genet. Cytol. 24:227-233.
Islam, A.K.M.R. and K.W. Shepherd. 1981. Wheat-barley addition lines: their use in genetics and evolutionary studies of barley. In Barley Genetics IV (Proc. IVth Int. Barley Genetics Symp.):729-739.
Pickering, R.A. 1983a. The location of a gene for incompatibility between Hordeum vulgare L. and H. bulbosum L. Heredity 51:455-459.
Pickering, R.A. 1983b. The assessment of variation in two populations of Hordeum bulbosum L. for improving success rates in a doubled haploid barley programme. Euphytica 32:903-910.
Pickering, R.A. and P.W. Morgan. 1978. Rep. Welsh P1. Breed. Stn, for 1977:65-66.
Riley, R. and V. Chapman. 1967. The inheritance in wheat of crossability with rye. Genet. Res., Camb. 9:259-267.
Snape, J.W., V. Chapman, J. Moss, C.E. Blanchard and T.E. Miller. 1979. The crossabilities of wheat varieties with Hordeum bulbosum. Heredity 42:291-298.
Thomas, J.B., P.J. Kaltsikes and R.G. Anderson. 1981. Relation between wheat-rye crossability and seed set of common wheat after pollination with other species in the Hordeae. Euphytica 30:121-127.
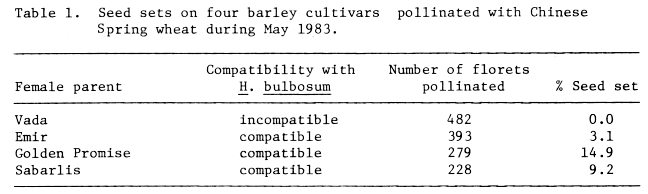{kind=link}
{kind=link}