II. 39. Reciprocal translocations in sodium azide treatments in barley.
A. R. Prina (1) G. Hagberg and A. Hagberg, Swedish University of Agricultural Sciences, Department of Plant Breeding, Svalov, Sweden.
(1)Fellow from the "Consejo Nacional de Investigaciones Cientificas y Tecnicas", Argentina.
Sodium azide is a very potent mutagen in barley and in other higher plant species (Kleinhofs et al. 1978). Its chromosome breakage effect in barley has been reported as absent or very low by many authors (Sideris 1968; Sideris et al. 1969, 1973; Sideris and Nilan 1970; Sander et al. 1972, 1978; Nilan et al. 1973, 1975; Nilan and Pearson 1975; Cortes 1973; Kleinhofs et al. 1974; Walther 1975; Drina and Favret 1983a) and the same has been found in Vicia (Kihlman 1959), soybean (Vig 1973), and human leucocytes (Nilan et al. 1975; Sander et al. 1978). On the other hand, it has been reported that in barley a high level of sterility in the M1 plants is induced by sodium azide (Cortes 1973; Prina 1978; Prina and Favret 1981, 1983a,b). Only one reciprocal translocation obtained in sodium azide treatments has been previously reported. This was shown to involve chromosomes 6 and 7 of normal barley (Lehman and Hagberg 1977).
In order to study the sodium azide mutachromosome effect and later on its possible specificity through reciprocal translocations, two different experiments were carried out. In one of them other meiotic abnormalities, chlorophyll mutants, and sterility of the M2 plants were also measured.
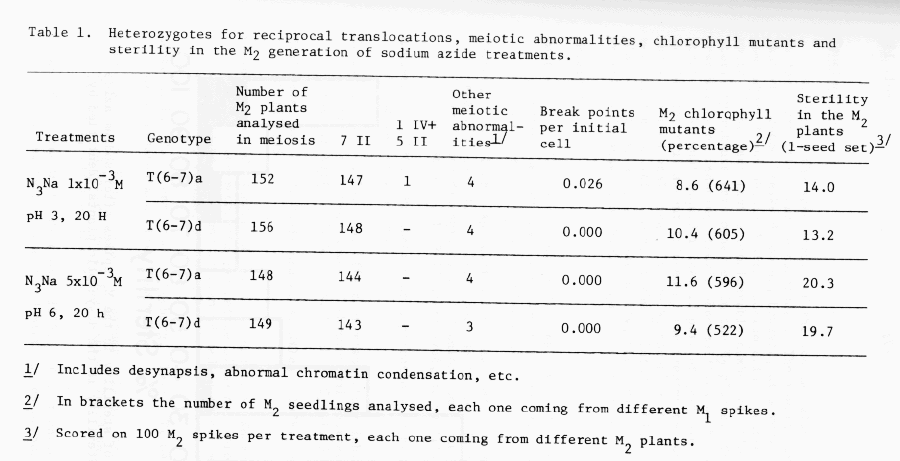
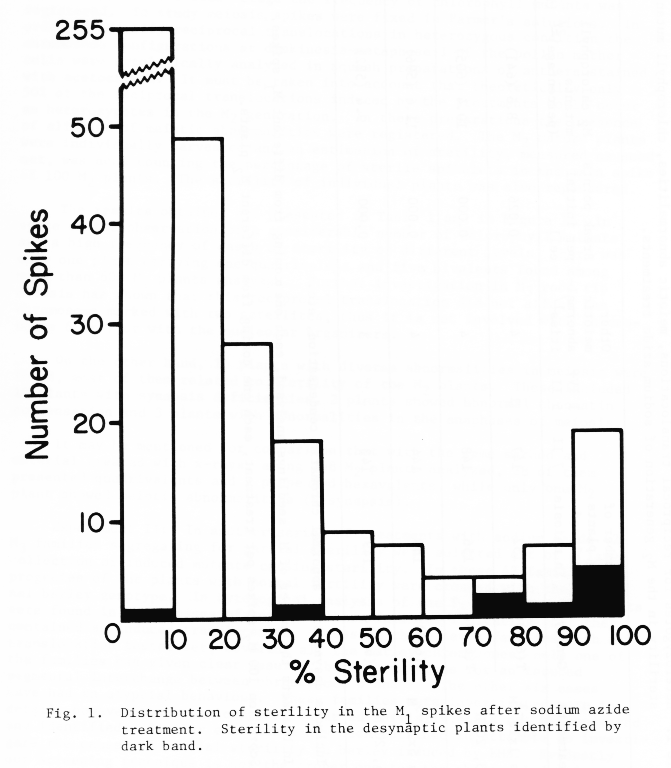
Experiment I: Two translocation lines (T6-7a and T6-7d) were used which have the two satellites in opposite arms of one of the translocated chromosomes (Hagberg 1962). It would allow easily to detect studying the positions of the satellites in root tips cytological preparations whether this particular marker chromosome is affected by the reciprocal translocations or not (Favret 1968). Seeds of the two translocation lines were treated with sodium azide (1 x 10-3M, pH 6 and 5 x 10-3M, pH 3, during 20 h) and immediately sown in the field nursery. Four spikes per M1 plant were harvested and one seed from each spike was bulked for each treatment. This M2 population was also sown in the field nursery and when the seedlings were in the "second leaf" stage the frequency of chlorophyll mutants was registered. To study meiosis, spikes were fixed in Farmer's solutions. In order to identify reciprocal translocations in heterozygous condition the chromosome configurations at diakinesis-metaphase I of the pollen mother cells were cytologically analysed in squash preparations of anthers stained with acetocarmin. It must be taken into account that theoretically only 50% of the reciprocal translocations induced by the treatments would occur as heterozygotes in the M2 generation. In these preparations the presence of all kind of meiotic abnormalities were registered. The M2 mature plants were individually harvested and an estimation of sterility, measured as seed set, was done counting the percentage of sterile spikelets in the main spike of 100 M2 plants. The sterility of individual plants was also registered.
The results obtained are presented in Table 1 and in Figure 1. In spite of the observation of a considerable number of chlorophyll mutants and a high frequency of cases of sterility of different levels, there was only one plant carrying one quadrivalent and five bivalents found among more than 600 M2 plants analyzed. Further investigation in M3 root tip mitosis has shown that this reciprocal translocation did not affect the chromosome marked with two satellites, thus it is not involved with chromosomes 6 or 7 or with the nucleolar organizers.
On the other hand, 15 plants with diverse abnormalities in meiosis were found, most of them related to sterility of the M2 plants. These included 10 plants with synapsis deficiencies, 2 plants showed abnormal chromatin condensation and 3 plants with abnormalities in the anaphases of meiosis.
It may be mentioned for comparison that with the same method and material treated with x-rays, among 368 M2 plants analyzed, 28 of them presented quadrivalents and 2 presented hexavalents, while only one M2 plant showed meiotic abnormalities (desynapsis).
Experiment II:
In another series of treatments with sodium azide 42 M3 families segregating
for partial sterility were selected out of a large collection of induced
mutants causing sterility. In these 42 families the progenies of the plants
with normal fertility were crossed with the original barley genotype. In
cytological analysis of the F1 spikes quadrivalents were found in the crosses
of seven of these families indicating that they contained an induced translocation.
Crossing these seven families with a translocation tester set, covering
all the barley chromosomes, only one of the families has given clear results,
being homozygous for an induced segmental interchange between chromosomes
6 and 7. The other six cases have had an atypcial behaviour. These families
were investigated more in detail, however the translocations observed initially
could not be recovered and identified. Similar experience has been obtained
in attempts to investigate the causes of partial sterility in barley induced
by EMS. Evidently our screening technique is not the proper one for the
changes induced.
Conclusions
Regarding the specificity of the sodium azide mutachromosome effect
so far, very little can be concluded. The results presented here confirm
that chromosome breaks do occur, however, at a very low frequency. Chromosome
aberrations obtained after sodium azide treatments are very rare in spite
of the fact that a considerable number of chlorophyll mutants and sterility
mutants were observed in the M2 plants. On the other hand, the possibility
of inducing meiotic abnormalities with sodium azide is demonstrated. We
must consider that if part of them are regulated by recessive alleles the
real amount induced by the treatments can be much higher than those observed
analysing only one M2 plant per M1 spike progeny. These meiotic abnormalities,
most of them related to high sterility in the M2 plants, can explain, at
least partly, the sodium azide induced genetic sterility in absence of
high frequencies of visible chromosome aberrations.
References:
Cortes, H. 1973. Mutaciones inducidas por azida s6dica y rayos X en
tratamientos simples y combinados en cebada. Tesis M.S. Esc. Grad. Ciencias
Agrop. INTA, Castelar/Argentina.
Favret, E. A. 1968. Aplicaciones de las radiaciones ionizantes en la Genetica Vegetal. A. licaciones de la energia atomica al aumento de la productividad agricola. I Reuni6n Com. Interamericana Energia Nuclear. Stgo. Chile 9-12 Enero 1968, p. 95-111.
Hagberg, A. 1962. Production of duplications in barley breeding. Hereditas 48:243-246.
Kihlman, B. A. 1959. The effect of respiratory inhibitors and chelating agents on the frequencies of chromosomal aberrations produced by x-rays in Vicia. J. Biophys. Biochem. Cytol. 5:479-490.
Kleinhofs, A., W. M. Owais and R. A. Nilan. 1978. Azide. Mutat. Res. 55:165-195.
Kleinhofs, A., C. Sander, R. A. Nilan and C. F. Konzak. 1974. Azide mutagenecity-mechanism and nature of mutants produced. In Poliploidy and Induced Mutations in Plant Breeding (Proc. meeting Bari, FAO/IAEA, Oct. 1972, Vienna, p. 195-199.
Lehmann, L., P. Hagberg and G. Hagberg. 1977. Translocations in sodium azide treated material. BGN 7:43.
A. Kleinhofs and C. Sander. 1975. Azide mutagenesis in barley. In Barley Genetics III (Proc. Third Barley Genetics Symp., Garching, 1975) p. 113-122.
Nilan, R. A. and O. W. Pearson. 1975. Lack of chromosome breakage by azide in embryonic shoots and microspores of barley. BGN 5:33.
Nilan, R. A., E. G. Sideris, A. Kleinhofs, C. Sander and C. F. Konzak. 1973. Azide - a potent mutagen. Mutat. Res. 17:142-144.
Prina, A. R. 1978. Efecto mutagenico y mutacromos6mico de la azida sodica en cebada. Tesis M.S. Esc. Grad. Ciencias Agrop. INTA, Castelar, Argentina.
Prina, A. R. and E. A. Favret. 1981. Comparative analysis of the somatic mutation process in barley. In Barley Genetics IV (Proc. 4th Barley Gen. Symp., Edinburgh 1981): p. 886-891.
Prina, A. R. and E. A. Favret. 1983a Parabolic effect in sodium azide mutagenesis in barley. Hereditas 98: 1. In press.
Prina, A. R. and E. A. Favret. 1983b. Influence of potassium cyanide on the azide mutagenesis in barley. Hereditas 98: 2. In press.
Sander, C., A. Kleinhofs, C. Konzak and R. A. Nilan. 1972. Kinetic studies on the induction of the inhibition of repair of radiation damage in barley. BGN 2:71.
Sander, C., R. A. Nilan, A. Kleinhofs and B. K. Vig. 1978. Mutagenic and chromosome-breaking effects of azide in barley and human leucocytes. Mutat. Res. 50:67-75.
Sideris, E. G. 1968. Physiological and cytogenetical aspects of the effect of sodium azide on irradiated and non-irradiated barley seeds. Ph.D. thesis, Washington State University, Pullman, USA.
Sideris, E. G. and R. A. Nilan. 1970. The effect of a metabolic inhibitor on the frequency of radiation-induced chromosome aberrations and mutations in Hordeum vulgare (abstr.). Rad. Res. 43:245-246.
Sideris, E. G., R. A. Nilan and T. P. Bogyo. 1973. Differential effect of sodium azide on the frequency of radiation-induced chromosome aberrations vs. the frequency of radiation-induced chlorophyll mutations in Hordeum vulgare. Radiat. Bot. 13:315-322.
Sideris, E. G., R. A. Nilan and C. F. Konzak. 1969. Relation of radiationinduced damage in barley seeds to the inhibition of certain oxidoreductases by sodium azide. In: Induced Mutations in Plants (Proc. Symp. Pullman, FA0/IAEA, July 1969) IAEA, Vienna, p. 313-321.
Vig. B. K. 1973. Somatic crossing-over in Glycine max (L) Merril. Mutagenicity of sodium azide and lack of sinergistic effect with caffeine and mytomycin C. Genetics 75:265-277.
Walther, F. 1975. The influence of storage on sodium azide treated barley seeds and on the efficiency of the chemomutagen. In Barley Genetics III (Proc. 3rd Barley Gen. Symp., Garching 1975) p. 123-131.
{kind=link}
{kind=link}