II. 2. The hap gene causes facultative pseudogamy in barley.
R. A. Finch, Plant Breeding Institute, Maris Lane, Trumpington, Cambridge, England. R
Embryo sac and seed development were studied in Feulgen-stained squash preparations of 3 homozygous hap lines (without tig-034 or let) kindly supplied by Professor A. Hagberg, who derived them from the Bonus tig-034 mutant (Hagberg and Hagberg, 1980). The lines were: hap 81/1154-614-2 from the F4 of Mona x Bonus mutant; VH 81/1146-128-1(44)C3 from a colchicine-doubled haploid of Mona x Bonus mutant; and hap D4-C2 from a colchicine-doubled haploid line of the Bonus mutant (A. Hagberg, pers. comm.).
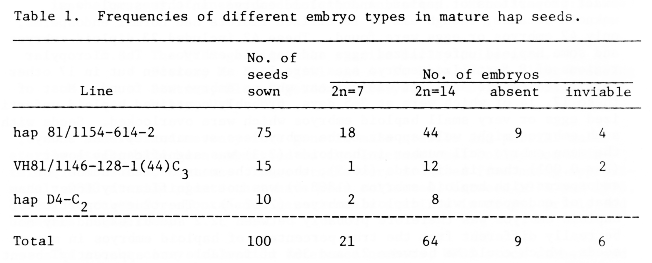
All seeds looked normal, but embryos in some were haploid, inviable or absent (Table 1). Lines did not differ significantly in frequencies of different seed types. When left to self in glasshouses at Cambridge, all lines varied from nearly normal seedset to over 50% failure of seed formation in different spikes, when normal barleys showed nearly full fertility. Both male and female fertility can be high, however, as hand pollinating emasculated hap 81/1154-614-2 spikes in a growth room at 20°C in continuous light gave fertilization rates of up to 85.2% for pollen from the same line.
Table 1. Frequencies of different embryo types in mature hap seeds.
In glasshouse grown VH 81/1146-128-1(44)C3, no abnormality in female development was seen at pachytene (2 cells), metaphase I and anaphase II (1 cell each) or in 26 florets with embryo sacs at a range of stages between meiosis and readiness for pollination. Meiosis, tetrads and first pollen grain mitosis were normal in anthers. In aceto-carmine squashes, 98.6% of 1050 mature pollen grains (50 or 100 grains from each of 13 anthers) had 2 male gametes each and looked normal.
Seed development was studied in hap 81/1154-614-2 grown in the growth room. Unpollinated plants were not spontaneously parthenogenetic, for when 2 spikes (24 and 23 florets) were emasculated just before the dehiscence stage and left bagged and unpollinated for a week before fixation, no seed was set. Squashes showed that all 47 florets had no embryo or endosperm.
Squashes of 147 florets from 8 emasculated spikes fixed at 24-120 h after hand pollination with hap pollen showed that 67 (45.6%) were fertilized. In 1 floret 2 days after pollination, the embryo consisted of 2 cells, both with a prophase nucleus where 7 chromosomes could be counted. This embryo was accompanied by 2 synergids and a normal endosperm with 1795 interphase nuclei. One synergid was intact and the other contained its own degenerated nucleus, granules from the pollen tube and what was possibly a sperm nucleus still completely condensed. There was no sign of chromosome elimination. In 3 florets at 3 days, an egg cell (clearly recognizable by its characteristic cytoplasmic granules and large diffuse pale-stained haploid interphase nucleus) was accompanied by a large normal endosperm with mitoses where 21 chromosomes could be counted. In a floret at 5 days, an embryo with 39 cells including some at mitosis where 2n=7, the large endosperm had several mitoses with 21 chromosomes. Embryos with diploid mitoses were also accompanied by endosperms with triploid mitoses in several florets. In many florets there were no endosperm mitoses and so the endosperm chromosome number could not be determined. All such endosperms looked normal and there was no evidence that any was not triploid. Many embryos also lacked mitoses, but haploid embryos were easily identified by their obviously low nuclear DNA amounts, compared with diploid ones. The exact proportions of haploid and diploid embryos in this sample was unknown because, though all 67 fertilized florets had normal endosperms, only 42 embryos or eggs were found. These 42 included 23 diploid embryos and some haploid unfertilized eggs and haploid embryos. The micropylar regions of 8 fertilized embryo sacs were lost at excision but in 17 other fertilized embryo sacs, it was unclear why no embryo was found. Most of these 17, 14 of which were at 3-5 days, probably contained haploid unfertilized eggs or very small haploid embryos which were overlooked. Seeds with such embryos might well appear to be embryoless at maturity. At 2 days, the mean embryo cell number in haploids (2.3) was significantly lower (P < 0.001) than in diploids (16.5), though the mean nuclear number of endosperms with haploid embryos (1868.5) was not significantly lower than that of endosperms with diploid embryos (2012.7). The true proportion of haploid embryos was therefore probably between 32.2 and 61.0% and may not be really different from the true percentage of haploid embryos in mature seeds, which could be between 21 and 36% if inviable and apparently absent embryos are counted as haploid (Table 1).
At 24 h, the nuclear numbers in diploid embryos and 10 of 11 endosperms were within the known ranges of normal barley in our conditions and since haploid embryos tended to be smaller than diploid ones, it is most unlikely that embryo or endosperm development started before fertilization. Many unfertilized florets may have been unfertilized because too little pollen was used. A normal mature egg, pair of intact synergids and a polar nucleus were found complete in 43 unfertilized florets and were probably lost in dissection of many others where less than a complete set was found. There was no evidence that embryos developed from any cell type other than the fertilized or unfertilized egg. Haploid formation is clearly caused by development of unfertilized eggs after normal endosperm fertilization. Apomixis in hap plants is thus by facultative pseudogamy and haploid parthenogenesis.
In florets with fertilized endosperms, some unfertilized eggs had a Feulgen-positive, sperm-like body beside them in squash preparations. The identity of such bodies was uncertain, however, and it was impossible to be sure whether one or both sperms entered the embryo sac.
The 2 components of double fertilization, fertilization of the egg and fertilization of the polar nucleus, are clearly separable and hap should prove useful in studies of the mechanism of fertilization. The ability of hap eggs to develop parthenogenetically may also hold out the possibility of apomictic barley breeding (Hagberg and Hagberg, 1980). Crosses have been made at Cambridge to try to combine hap with tri stocks (Finch and Bennett, 1979) so that parthenogenetic diploids rather than haploids can be produced.
References:
Finch, R. A. and M. D. Bennett. 1979. Action of triploid inducer (tri) on meiosis in barley (Hordeum vulgare L.). Heredity 43:87-93.
Hagberg, A. and G. Hagberg. 1980. High frequency of spontaneous haploids in the progeny of an induced mutation in barley. Hereditas 93:341-343.
{kind=link}