II. 22. Back mutation of multiple mutants in the cer-cqu region.
Søgaard, B. and Wettstein-Knowles, P. von, Department of Physiology, Carlsberg Laboratory, G1. Carlsberg Vej 10, DK-2500 Copenhagen Valby, and Institute of Genetics, University of Copenhagen, Øster Farmigasgade 2A, DK-1353 Copenhagen K, Denmark "R".
Among 1252 allele-tested cer mutants, 13 were found to be multiple events. Only cer-c, -q and -u wem involved, and all possible combinations were identified. These 13 multiple mutations represent 3% of the total mutants of cer-c, -q and -u. Eleven of them were induced by neutrons, one by X-rays and one by ethyl methanesulfonate. While genetic tests failed to indicate the linear order of cer-c, -q and -u on chromosome 4, they did demonstrate that the distance between any pair apparently was less than 0.025 map units (Wettstein-Knowles, 1979; Wettstein-Knowles and Søgaard, 1980). Cer-c, -q and -u are known to act in the common biochemical pathway giving rise to the beta-diketones, hydroxy-beta-diketones and esterified alkan-2-ols which are prominent components of the epicuticular wax on the uppermost leaf sheaths and internodes as well as given parts of the spike (see Wettstein-Knowles, 1979; Mikkelsen, 1979).
Two interpretations of the above observations are feasible. Either the cer-cqu region comprises three closely linked genes, i.e., a gene-cluster, or it is a single gene coding for a polypeptide chain with three functional domains, i.e., a cluster-gene or multifunctional-gene. In the former case the multiple mutants have to comprise large deletions and separate mutational changes. In the latter case a single mutational change, e.g. of a base-pair, can effect one, two or all three of the functional domains of the polypeptide, resulting in the single, double and triple phenotypes. In a multifunctional-gene (but not in a gene-cluster) single back mutations which revert double and triple phenotypes to wild type should occur at a reasonable frequency.
In the summer of 1978 cer-cqu multiple mutants were crossed to cer-j59 on chromosome 4. the Fl's were grown in the phytotron at Oslo, Norway during the following winter, and the F2's planted at Risø, Denmark in the summer of 1979. Seeds homozygous for cer-cqu and cer-j were multiplied at Christ Church, New Zealand during the winter. In the spring of 1980 seeds from four of the cer-cqu multiple mutants marked with cer-j were treated with NaN3. Since all four cer phenotypes are recessive to the wild type, back mutations will result in wild type amounts of wax on either the leaf sheaths, internodes and spikes (cer-cqu) or leaves (cer-j). Back mutations are expected to yield in the M1 chimeric plants, some shoots or parts of shoots being of wild type, others of mutant phenotype. The cer-j59 marker was included to allow us to discriminate between reversions and unwanted cross pollinations by stray pollen or inmixtures during seed increase as well as after NaN3 treatment.
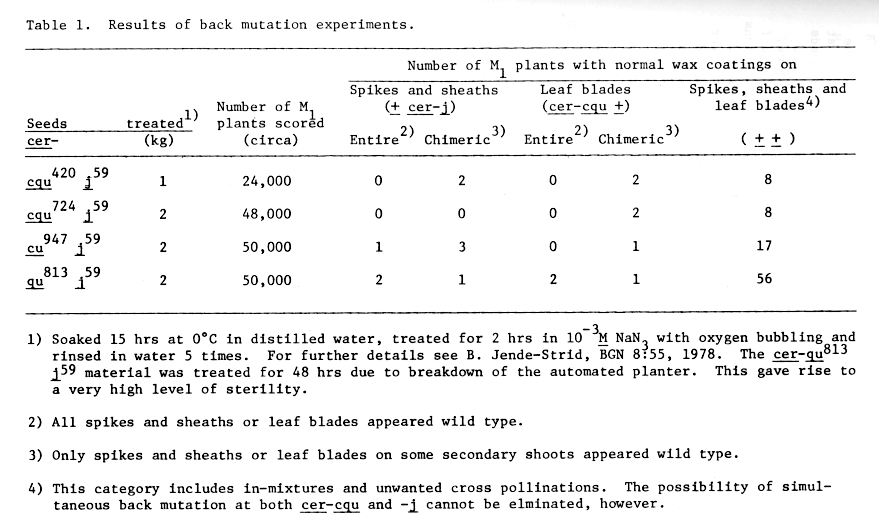
Table 1 summarizes the results of the first back mutation experiments. Nine apparent back mutation events at cer-cqu (columns 4 and 5) and eight at cer-j (columns 6 and 7) were identified among the M1 plants. Whereas cer-j59 appears to back mutate when combined with all four of the cer-cqu multiple mutants, only three of the latter back mutated, namely, cer-cqu420, -cu947 and -qu813. Before we conclude that cer-cqu724 cannot be back mutated, additional tests will be made.
Table 1. Results of back mutation experiments.
Not included in Table 1 are two additional interesting plants on which the amount of wax visible was intermediate between that of the mutant (none) and the wild type. That is, among the M1 plants of the cer-qu813 -j59 NaN3 treated seeds were two plants that had an intermediate amount of wax on the stems and spikes. The leaf blades of one of these was missing wax (cer-j59 in phenotype) while those of the other had wild type amounts of wax.
Seeds were collected from all M1 plants in which the cer phenotype was altered to wild type or an intermediate amount of wax on any organ. From the chimeric plants each spike was harvested separately. To verify the M1 phenotypes we are growing the M2 plants in the phytotron at Stockhom, Sweden. Thus far we have confirmed on the M2 plants the M1 phenotypes of all 9 cer-cqu and 8 cer-j reversions to wild type. Crosses will be carried out to determine whether the back mutational events are the result of mutations within the cer-cqu or cer-j regions themselves or result from suppressor mutations.
On the basis of our present M1 and M2 results, we conclude that cer-cqu420 -cu947 and -qu813 each resulted from single mutational events affecting 3, 2 and 2 different domains of the multifunctional polypeptide determined by the cluster-gene cer-cqu.
Acknowledgments:
We thank Barbro Jende-Strid for making the NaN3 treatments and Diter v. Wettstein for providing a bottle of champagne for each identified cer-cqu reversion.
References:
Mikkelsen, J. D. 1979. Structure and biosynthesis of beta-diketones in barley spike epicuticular wax. Carlsberg Res. Commun. 44:133-147.
Wettstein-Knowles, P. von. 1979. Genetics and biosynthesis of plant epicuticular waxes. In: Biochemistry and physiology of Plant Lipids. Appelqvist, L. A. and Liljenberg, C. Eds. Elsevier/North Holland Biomedical Press, Amsterdam. pp. 1-26.
Wettstein-Knowles, P. von and B. Søgaard. 1980. The cer-cqu region in barley: gene cluster or multifunctional gene. Carlsberg Res. Commun. 45:125-141.
{kind=link}