II. 20. Related response of barley cultivars to the "bulbosum-" and anther-culture techniques of haploid production.
Bärbel Foroughi-Wehr (l), Richard Pickering (2) and Wolfgang Friedt (l). (l)Biologische Bundesanstalt fur Land- und Forstwirtschaft, Institut fur Resistenzgenetik, D-8059 Grünbach, W.Germany. (2)Welsh Plant Breeding Station, Plas Gogerddan near Aberystwyth, SY23-3EB, Great Britain.
It has been demonstrated in a number of investigations that seed set and embryo differentiation in crosses of Hordeum vulgare x H. bulbosum are under the control of both parental genotypes (Jensen 1976; Novak et al. 1977; Pickering 1980). It was also clearly demonstrated by Pickering and Morgan (1979) that not only the genotype of H. bulbosum but also that of H. vulgare influence chromosome elimination and hence the proportion of haploid plants produced.
Earlier work on anther culture of barley (H. vulgare) also indicated genotypic influence on response of anther culture in vitro. Foroughi-Wehr et al. (1976) succeeded in inducing callus formation from anthers of twenty spring-barley cultivars. Striking differences between varieties in the proportion of callus-producing anthers and regenerated plants were recorded.
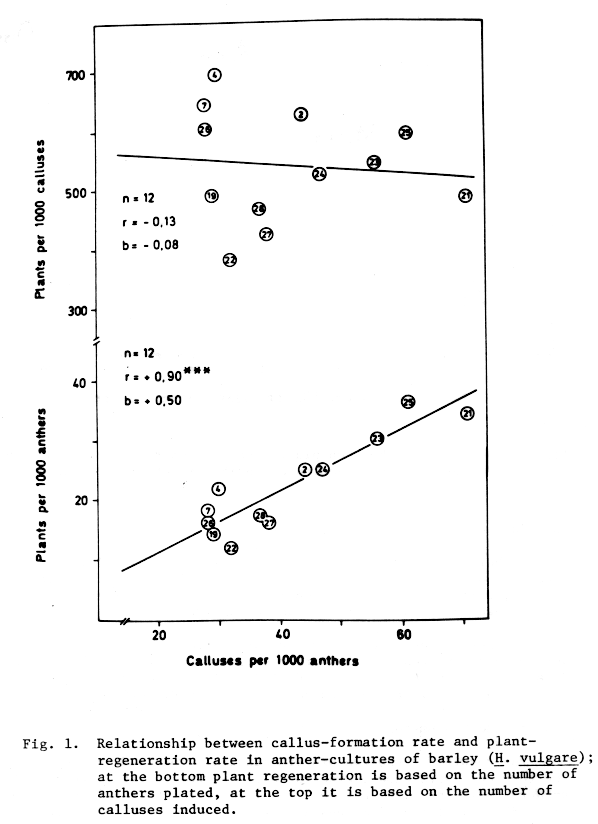
These previous findings of genotypic differences in response to anther culture have been confirmed in more recent work. When anthers of more than 30 different spring-barley crosses have been cultured during 1979 the proportion of callusing varied between 5 to 73%. From 0.3 to 4.8% of the anthers sown gave rise to plants (Friedt and Foroughi-Wehr 1980). The respective data of 12 crosses with the cultivar 'Trumph' are presented in Fig. 1. Callus formation and plant regeneration are closely related when both criteria are based on the number of cultured anthers (Fig. 1 bottom). However, when the number of regenerated plants is based on the number of induced calluses, then no relationship between callus formation and plant regeneration can be detected (Fig. 1 above). This can be interpreted such that both features are independent phenomena. Therefore, varieties which are highly productive with regard to callusing do not necessarily yield maximum numbers of plants (see e.g. cross No. 21 in Fig. 1). The different response of crosses to anther culture are obviously due to the genotypes of the respective second parents. Although genotypic influences were detected in separate experiments on embryo- and anther-culture, no information was available up to now regarding the response of the same genotypes to both methods of haploid production. The present study attempted to find out the response of different barley cultivars to the two methods.
Materials and Methods:
Anther donor plants of the spring barley cultivars 'Amsel', 'Dissa', 'Edelmut' and 'Mutina' were cultivated under field conditions at Grünbach, Bavaria, during the growing season 1975. The method of anther culture and plant regeneration are described in Foroughi-Wehr et al. (1976).
Crosses between the four H. vulgare varieties and H. bulbosum stocks were made during February 1980 in the glasshouse at the Welsh Plant Breeding Station, Aberystwyth. Full details of the environmental conditions and doubled haploid techniques may be found in Pickering (1980).
Results and Discussion:
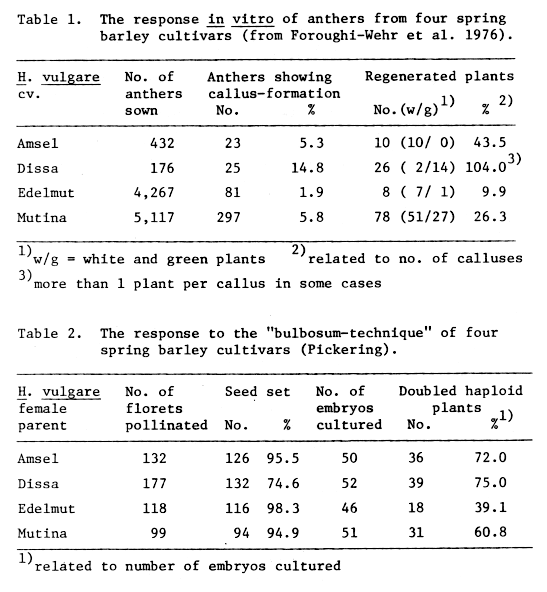
The results of the two experiments are summarized in Tables 1 and 2. The four cultivars show a wide range of figures for the percentages of anthers callusing with 'Edelmut' the lowest at 1.9% and 'Dissa' the highest at 14.8%. Similar differences appear for the rates of plant regeneration.
With 'Dissa' a total of 26 plants were regenerated out of only 176 anthers plated (Foroughi-Wehr et al. 1976), which is a plant regeneration rate of 9%.
Table 1. The response in vitro of anthers from four
spring barley cultivars (from Foroughi-Wehr et al. 1976).
Table 2. The response to the "bulbosum-technique"
of four spring barley cultivars (Pickering).
In the "bulbosum-experiment" the proportion of cultured embryos varied according to the cultivars used as the female parent (Table 2). Despite culturing similarly differentiated embryos, differences between the varieties were recorded for doubled haploid plant production. Such variation has been shown to be solely under the control of the H. vulgare parent (Simpson et al. 1980). Although embryo dormancy has been observed in Avena fatua (Naylor and Simpson 1961) very few instances of its occurrence have been reported in H. vulgare (Norstog and Klein 1972). Nevertheless it was decided to determine whether it existed in a sample of freshly harvested barley (cv. 'Golden Promise') known to possess dormancy. It was found that lemma removal and excision of the distal portion of the caryopsis resulted in high levels of germination and thus no embryo dormancy was shown to exist (Pickering, 1980).
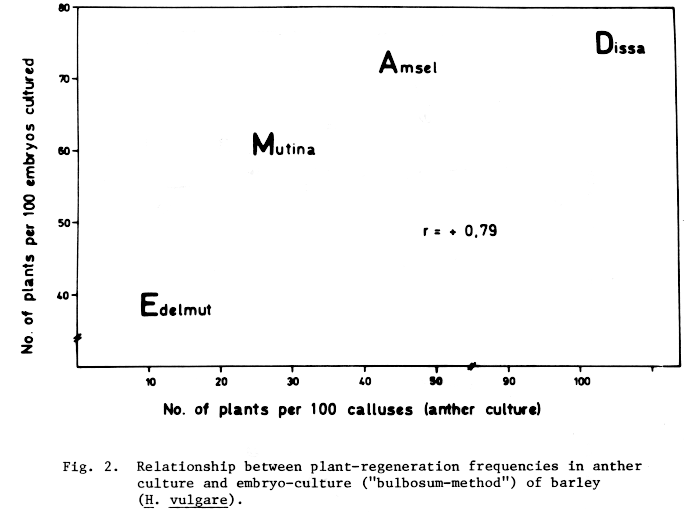
In Figure 2 the response in both experiments is presented diagrammatically. It is evident that rates of regenerated plants in the two methods are rather closely related (r = +0.79). Such a relationship is not to be expected a priori, since in either method different plant parts in different developmental states were cultured. Responsiveness of certain genotypes in tissue culture seems to be a general ability expressed even in different culture methods. Parallel although genotypic variation is observed in either method studied here, the range between genotypes (cultivars) in anther culture is much broader (10-100%) than in the bulbosum-method (40-75%). These genotypic differences are a major disadvantage for the practical application of haploid barley techniques, since not all of the valuable genotypes for plant breeding can be used in such unconventional methods.
There is no information available yet with regard to the genetic base of differential varietal response in tissue culture. Such information is needed for estimating the ease or difficulty of transferring the character "tissue-culture responsiveness" from one cultivar to another. Studies in this respect have been initiated.
References:
Foroughi-Wehr, B., C. Mix, H. Gaul and H. M. Wilson. 1976. Plant production from cultured anthers of Hordeum vulgare L. Z. Pflanzenzuchtg. 77:198-204.
Friedt, A. and B. Foroughi-Wehr. 1980. Moglichkeiten and Grenzen des Einsatzes von Zellkulturen in der Resistenzzuchtung bei Getreide. 31. Arbeitstagung der Saatzuchtleiter, Gumpenstein, 25-27.11.80 (in press).
Jensen, C. J. 1976. Barley monoploids and doubled monoploids: techniques and experience. In: Barley Genetics III, pp. 316-345, Proc. 3rd Int. Barley Genetics Symp., Munchen, 1975.
Naylor, J. M. and G. M. Simpson. 1961. Dormancy studies in seed of Avena fatua. 2. A gibberellin-sensitive inhibitory mechanism in the embryo. Can. J. Botany 39:281-295.
Norstog, K. and R. M. Klein. 1972. Development of cultured barley embryos. II. Precocious germination and dormancy. Can. J. Botany 50:1887-1894.
Novak, F. J., F. Minarik, L. Ohnoutkova and J. Bouma. 1977. Productior of haploid barley (Hordeum vulgare L.) and wheat (Triticum aestivum L.) in interspecific hybridization with Hordeum bulbosum L. In: Use of Tissue Cultures in Plant Breeding, pp. 427-439, Proc. Int. Symp., Olomouc, CSSR, Czechoslovak Aca. Sci., Prag.
Pickering, R. A. 1980. Use of the doubled haploid technique in barley breeding at the Welsh Plant Breeding Station. Rep. Welsh Plant Breed. Stn. for 1979, pp. 208-226.
Simpson, E., J. W. Snape and R. A. Finch. 1980. Variation between Hordeum bulbosum genotypes in their ability to produce haploids of barley, Hordeum vulgare. Z. Pflanzenzuchtg. 85, 205-211.
{kind=link}
{kind=link}
{kind=link}