Genetics and Mapping of Powdery Mildew Resistance in the Harrington/Chebec and Clipper/Sahara Doubled Haploid Populations
Brian Steffenson1, Peter Langridge2, and Roger
Effertz1
1Department of Plant Pathology
North Dakota State University, Fargo
and
2Department of Plant Science
University of Adelaide, Glen Osmond
Introduction. Parents of the Australian Barley Genome Mapping populations were found to exhibit differential reactions to isolate ND97GH of the powdery mildew pathogen (Blumeria graminis f. sp. hordei) in a preliminary study. Harrington and Clipper were resistant to this isolate, whereas Chebec and Sahara were susceptible. Two doubled haploid populations consisting of 56 and 60 individuals each from the crosses Harrington/Chebec and Clipper/Sahara, respectively, were evaluated to isolate ND97GH in the greenhouse. A Restriction Fragment Length Polymorphism (RFLP) marker map exists for both of these populations and was available from the Graingenes web site (Langridge et al. 1996). Disease reaction data of progeny from the two doubled haploid populations were merged with the RFLP marker data to map the resistance locus in each population.
Materials and Methods. Parents and doubled haploid progeny of the two populations were grown in 10 cm pots filled with a peat moss:perlite (3:1 ratio) potting mixture and grown at 21-23° C in a greenhouse where supplemental lighting was provided for 14 hr/day. Fertilizer was applied at planting with water soluble and controlled release formulations. Plants were inoculated by shaking conidia from heavily infected Larker barley plants onto the parents and doubled haploid progeny of both populations when the first leaves of the plants were fully expanded (eight days after planting). Isolate ND97GH exhibits a low virulence/high virulence pattern of P01, P02, P03, P04A, P04B, P06, P07, P08A, P08B, P09, P10, P11, P13, P16, P17, P19, P20, P21/P12, P14, P15, P18, P23, P24 on the Pallas differential genotypes of Kølster et al. (1986). The doubled haploid populations and parents were incubated under ambient greenhouse conditions (21-23° C). Infection types were assessed on the progeny and parents of both populations six and seven days after inoculation using the 0-4 rating scale of Torp et al. (1978). Infection types of 0, 1, and 2 were considered to be indicative of host resistance, whereas infection types of 3 and 4 were considered to be indicative of host susceptibility. Linkage analyses were performed and map positions of the resistance genes were determined using the MAPMAKER computer program (Lander et al.1987).
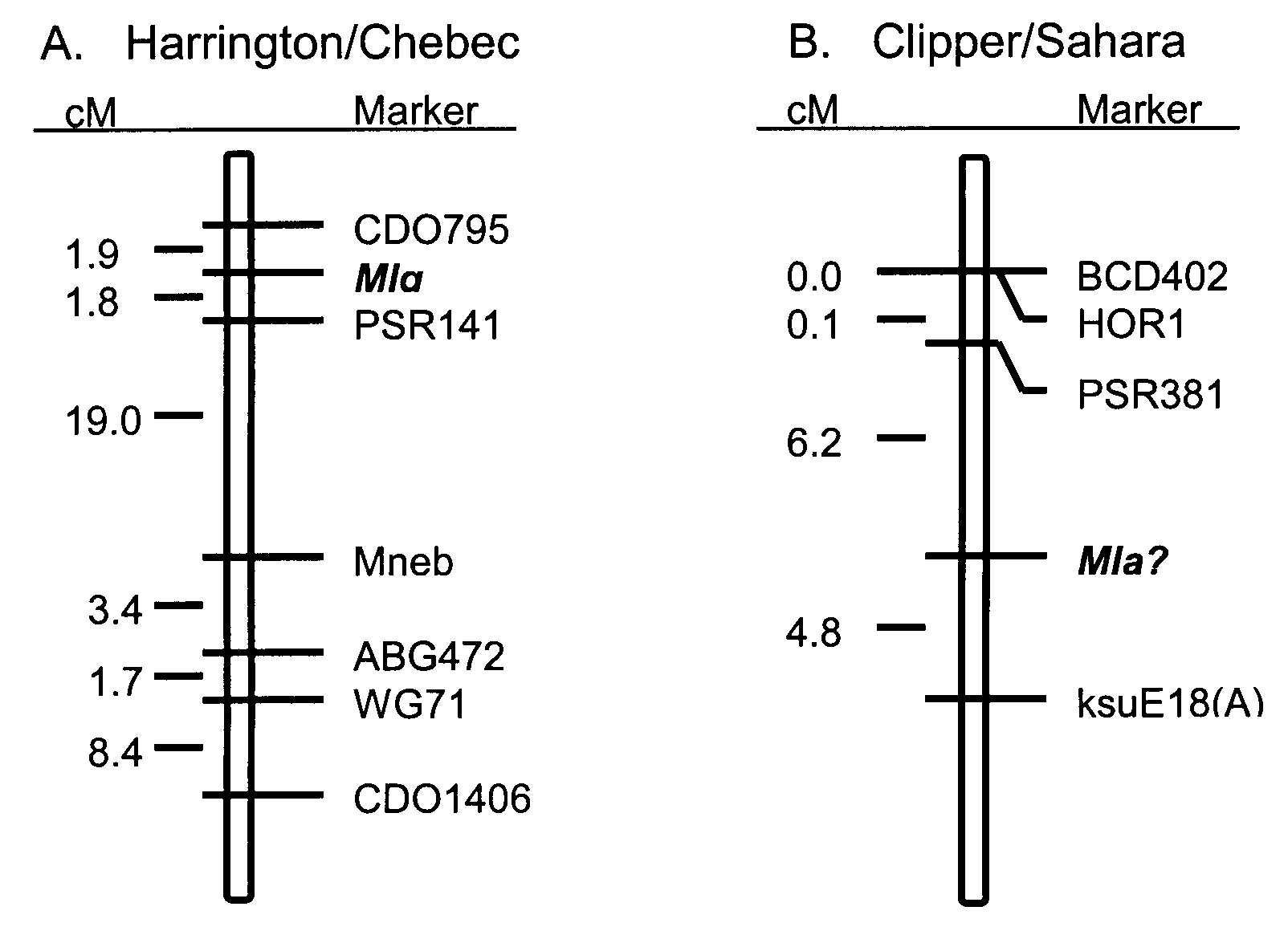
Results and Discussion. The infection types observed on the parents of the two populations were very distinct with Harrington exhibiting infection type 0, Clipper exhibiting infection type 0,1n (necrosis), Chebec exhibiting infection type 4, and Sahara exhibiting infection type 3,4. Infection types of the doubled haploid progeny of both populations were equally distinct, which allowed for the unequivocal classification of progeny into resistant and susceptible groups. The Harrington/Chebec and Clipper/Sahara populations segregated approximately 1:1 for resistance:susceptibility to isolate ND97GH of B. g. f. sp. hordei (Table 1), indicating the involvement of a single gene for resistance in each of these populations. The gene in the Harrington/Chebec population was mapped to chromosome 4(4H), and the gene in the Clipper/Sahara population was mapped to chromosome 5(1H). The powdery mildew resistance locus on chromosome 4(4H) was flanked by the RFLP markers CDO795 and PSR141 at 1.9 and 1.8 cM, respectively (Figure 1A). The powdery mildew resistance locus on chromosome 5(1H) was flanked by the RFLP markers PSR381 and ksuE18(A) at 6.2 and 4.8 cM, respectively (Figure 1B). The resistance gene in the Harrington/Chebec population was contributed by Harrington and is presumed to be Mlg (Steffenson and Kleinhofs 1995). Mlg is located approximately 24 cM proximal from the RFLP marker ABG472 in three populations where Harrington is the donor parent (Steffenson and Kleinhofs 1995). The resistance gene in the Clipper/Sahara population was contributed by Clipper and may be one of the alleles at the Mla locus.
References.
Kølster, P. L., et al. 1986. Crop Sci. 26:903-907.
Lander et al. 1987. Genomics 1:174-181.
Langridge et al. 1996. Waite Agric. Res. Inst., Univ. Adelaide. http://wheat.pw.usda.gov/graingenes.html
Steffenson, B. J. and Kleinhofs, A. 1995. Barley Gen. Newsl. 39:89-91.
Torp et al. 1978. pp75-102 In: Royal Vet. and Agric. Univ. Copenhagen,
Denmark.
Table 1. Segregation of the Harrington/Chebec and Clipper/Sahara doubled haploid populations to isolate ND97GH of Blumeria graminis f. sp. hordei.
| Population |
|
|
|
| Harrington/Chebec |
|
|
|
| Clipper/Sahara |
|
|
|
Figure 1. Linkage maps of the powdery mildew resistance genes
with associated RFLP markers on A) barley chromosome 4(4H) from the Harrington/Chebec
doubled haploid population and on B) barley chromosome 5(1 H) from the
Clipper/Sahara doubled haploid population.